Zygochlamys delicatula (Hutton, 1873)
HUTTON, F. W. 1873. Catalogue of the Tertiary Mollusca and Echinodermata of New Zealand, in the collection of the Colonial Museum. Wellington, Government Printer, for Colonial Museum and Geological Survey Dept. xvi + 48 p. [p. 30]
1873 Pecten delicatula Hutton, 1873
1916 Chlamys subantarctica Hedley, 1916
1924 Chlamys campbellicus Odhner, 1924 [partim]
1925 Chlamys instar Iredale, 1925
1928 Chlamys titirangiensis Marwick, 1928
1916 Chlamys subantarctica Hedley, 1916
1924 Chlamys campbellicus Odhner, 1924 [partim]
1925 Chlamys instar Iredale, 1925
1928 Chlamys titirangiensis Marwick, 1928

Zygochlamys delicatula (Hutton); A. G. Beu, 1995, Pliocene Limestones and their scallops, figures 23b, c, e.
|
«101. P. delicatula, sp. nov. Sub-orbicular, rather compressed; with about twenty-seven narrow distant smooth rays, the interstices with three to five thin raised scaly lines; ears unequal, with radiating scaly ribs. Height, 2; length, 2.
Localities.— Castle Point, Wellington; Chatham Islands.» FREDERICK WOLLASTON HUTTON, 1873
|














|
«Description. Shell up to 110 mm high, most specimens smaller, to c. 70 mm; thin to more solid (Macquarie Island morph), adults almost circular, moderately inflated, left valve of most specimens a little more convex than right, equivalve, equilateral, anterior auricles larger and longer than posterior ones, umbonal angle of juveniles c. 85–90°, of adults c. 115–120°; most specimens white or yellowish, others orange, red or cream.
Both valves sculptured with c. 20 moderately prominent radial plicae; with narrow plicae with broader, evenly concave interspaces on left valve, wider plicae with narrower interspaces on right valve, subdivided and flanked by lamellose secondary and tertiary radial riblets. Antimarginal microsculpture in pre-radial stage and intercostally (Beu & Darragh, 2001: figs 41C, E; Dijkstra & Marshall, 2008: figs 53C, 54C). Auricles with 10–16 squamous radial riblets, except 4–5 riblets on anterior auricle of right valve. Byssal notch moderately deep, byssal fasciole broad, both decreasing in prominence with age. Functional ctenolium well-developed in juveniles, with 4–5 teeth, decreasing in number with age. Internal radial furrows present, edges not carinate. Hinge with moderately weak resilial and dorsal teeth. Dimensions. Illustrated specimen: VIC, E of Gabo I, 208–223 m (AM C.346889), large single lv: H 89.6, L 90.5 mm.
Habitat. Living in the sublittoral to bathyal zones, many specimens encrusted with sponges. However, not known living around Australia.
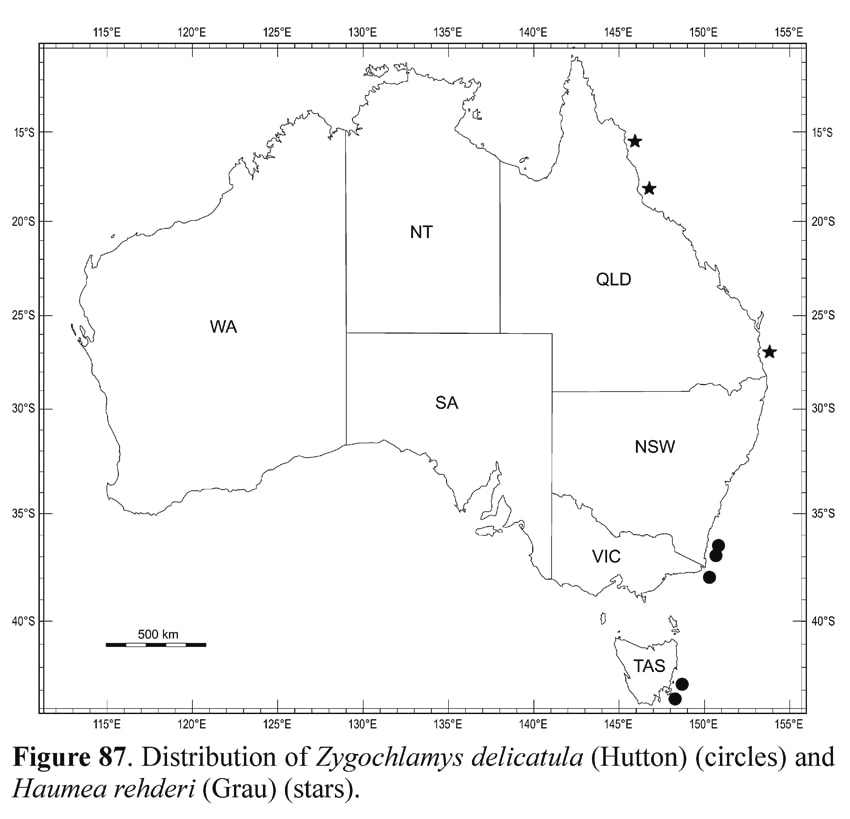
Distribution. Cook Strait to the southern region of the South Island, New Zealand, southwards to the Bounty, Antipodes, Auckland, Campbell and Macquarie Islands; dead specimens on the Chatham Rise, off W Auckland and N of the Three Kings Islands, New Zealand (Beu, 1999; Dijkstra & Marshall, 2008: fig. 56), westwards to southernmost New South Wales, eastern Victoria and eastern Tasmania. Present specimens dead at 140–282 m.
Remarks. The present specimens from Australia are Last Glacial Maximum fossils and no living records are known from Australia (see also Beu & Darragh, 2001: 121). Specimens from southeastern Australia have been incorrectly identified in the past as “Mimachlamys asperrima” or “Mimachlamys antiaustralis” by Hedley & May (1908), Hedley (1911) and May (1921) (Beu, 1985).
For information on biostratigraphy see Beu (1995: 16).» DIJKSTRA, H. H. & A. G. BEU. 2018. Living scallops of Australia and adjacent waters (Mollusca: Bivalvia: Pectinoidea: Propeamussiidae, Cyclochlamydidae and Pectinidae). Records of the Australian Museum, 70 (2): 113-330, figs. 1-102. [p. 274]
|

Zygochlamys delicatula (Hutton); H. H. Dijkstra & A. G. Beu, 2018, Living scallops of australia and adjacent waters, figure 82F.
|
|
«Chlamys/Zygochlamys delicatula has generally been interpreted as a variable species ranging from Late Pliocene to Recent (Dell 1964; Powell 1979; Waller 1991; Beu 1995, 1999; Beu and Darragh 2002), and for a time it was interpreted as a geographic subspecies of the southern South American Z. patagonica (King & Broderip, 1832) (Beu 1985b; Beu and Maxwell 1990). In the most recent study of the group, Jonkers (2003) introduced Psychrochlamys for Pecten patagonicus, and recognised two chronosubspecies of the Late Pliocene P. delicatula delicatula: P. delicatula subantarctica (Hedley, 1916) for the living forms, and P. delicatula instar (Iredale, 1925) for some subrecent valves from Cook Strait and off the Snares Islands (14C: 19,000 YBP; 18,850 6 650 YBP; 14,319 6 86 YBP) and Tasmania. Jonkers (2003, figs 34, 38) provided extensive statistical data showing differences between Recent specimens and the lectotype and other Late Pliocene specimens of P. delicatula that include smaller size (mean valve height 58.9 mm and 71.1 mm respectively), less symmetrical valves, less convex left valve, relatively shorter outer ligament, much lower auricular symmetry, angle between free margin of left valve anterior auricle and hinge <90° (versus >90°), much higher angle between free margin of posterior auricle and hinge (about 120° and 110° respectively), deeper byssal notch, and higher number of functional byssal teeth. Jonkers admitted that the New Zealand species identifed as P. delicatula instar were morphologically intermediate between P. delicatula delicatula and P. delicatula subantarctica, but differed in that both valves are less inflated than in either P. delicatula delicatula or P. delicatula subantarctica. It seems clear to us that Recent and subrecent forms are forms of a single species in which various shell parameters have undergone random drift over time. We prefer to interpret these subrecent shells as simply forms of Zygochlamys delicatula rather than chronosubspecies, being influenced by the extreme variation between living populations.
In this connection, we draw attention to the fact that most of the local recent populations of Zygochlamys delicatula have distinctive combinations of shell sculpture, suggesting that the aggregations are largely self recruiting (in the absence of imperical data, Michael and Cranfield (2001) used data from “similar” pectinid species to infer length of larval life as 40–50 days for Z. delicatula). Specimens from Pukaki Rise (Figs 53, 54), all livetaken specimens from Antipodes Islands (Fig. 55B, E), all live-taken specimens from the Bounty Plateau with the exception of material from NIWA stations I712 (no sponge and normally sculptured) and A717 (1 spiny, sponge covered specimen among 37 ‘normally’ sculptured, unencrusted specimens in station A717), and one lot from off the Snares Islands (station D131), are particularly distinctive in having an unusually thin shell, with prominent, sharp spines on all of the radial ribs, including (and particularly) the summits of the primaries. As with Talochlamys dichroa (see above), many individuals tend to be distinctly one form or the other, but individuals commonly occur in which sculpture changes abruptly from spineless to spiny (Fig. 53E, F) (but not vice versa), the transition evidently being a response to settlement of sponge on the shell. Specimens without encrusting sponges from or adjacent to these stations are considerably thicker and spineless (Fig. 55A, D). The summits of the primary radial ribs are typically smooth in populations from elsewhere, whether or not there is sponge encrustation (Fig. 53E, F). This particular sponge (A717, D131) was identified as an undescribed species of Esperiopsis (Demospongiae: Poecilosclerida: Esperiopsidae) by Michelle Kelly (NIWA, Auckland). The thin, spiny form is almost indistinguishable from the consociata morph of Talochlamys dichroa (Suter, 1909) from off the Three Kings Islands, yet large size (height up to 79.5 mm), and the narrow outline of juveniles clearly indicates that they are forms of Z. delicatula rather than Z. dichroa (see below). This appears to be the first record of geographic variation in scallop dimorphism in response to sponge encrustation. Other examples of sponge-scallop mutualism have been discussed by Beu (1965), Forester (1979), Chernoff (1987), Pitcher and Butler (1987), and Pond (1992). The sponge will be discussed elsewhere by Michelle Kelly (in prep.). Recently, Marin and López Belluga (2005) have provided evidence that sponge coating on the bivalve Arca noae significantly reduces predation, and this seems likely to pertain in pectinids. Since both extremes of sculpture may occur on a single individual, and there is fluid intergradation in shell morphology within and between populations, it seems clear to us that a single highly variable species is involved.» DIJKSTRA, H. H. & B. A. MARSHALL. 2008. The recent Pectinoidea of the New Zealand region (Mollusca: Bivalvia: Propeamusiidae,Pectinidae and Spondylidae). Molluscan Research, 28 (1): 1-88, figs. 1-70. [p. 68]
|


Zygochlamys delicatula (Hutton, 1873); H. H. Dijkstra & B. A. Marshal, 2008, The recent Pectinoidea of the New Zealand region, figure 53: juvenile cleaned of thinly encrusting sponge; figure 54: juvenile naturally free of encrusting sponge.
|
|
«The specimens figured by Odhner (1924) as Chlamys campbellicus belong to two taxa: Beu (1995, p. 17) designated the specimen in Odhnert fig. 36 as the lectotype of C. campbellica (= Psychrochlamys delicatula subantarctica) and referred the two left valves in figs. 37 and 38 to T. dichroa. Odhner's fig. 39 shows sculptural details of the specimen in fig. 37.»
JONKERS, H. A. 2003. Late Cenozoic-Recent Pectinidae (Mollusca: Bivalvia) of the Southern Ocean and neighbouring regions. Monographs of Marine Mollusca, 5: i-viii + 1-125 pp, 17pls. Backhuys Publishers, Leiden [p. 56]
|

Chlamys campbellicus n. sp.; N. H. Odhner, 1924, Papers from Dr Th. Mortensen’s Pacific expedition, plate 2, figures 36-39.
|
|
«Stratigraphical age and distribution. Late Pliocene (early Nukumaruan; Fig. 25). The taxon is widespread in southeastern and eastern North Island, New Zealand; a list of all known collecting sites there was given by Beu (1995). Other records are from concretions dredged off northernmost North Island (Beu, 1999), and from norrhern South Island (A.G. Beu, pers. comm.200I; Fig.22). P delicatula delicatula occurs together with common Phialopecten triphooki (Zittel, 1864), locally common Towaipecten mariae (Finlay 1927), locally abundant Mesopeplum (Mesopeplum) convexum (Quoy & Gaimard, 1835), and Kaparachlamys? sp. (single specimen; Beu, 1995).
Remarks. Psychrochlamys delicatula delicatula differs morphometrically from P. delicatula subantarctica in its larger size (mean VH 71.1 mm and 58.9 mm, respectively), its much more symmetrical shells (Fig. 3B), a particularly more convex LV, a relatively longer outer ligament, much higher auricular symmetry, the angle berween the free margin of the LV anterior auricle and the hinge >90º (<90º in P. d. subantarctica), a much lower angle berween the free margin of the posterior auricle and the hinge (c. 110º and 120º, respectively), a shallower byssal notch, and a lower number of functional byssal teeth (Fig. 34). These differences are substantially greater than those between P. delicatula subantarctica and P. patagonica. There are qualitative differences too: radial sculpture of P. d. delicatula is often more robust than in P. d. subantarctica, with crests of ribs rather wider and mostly not scaly and the prominent dorsal hinge tooth of the modern taxon is usually not present in P. d. delicatula.» JONKERS, H. A. 2003. Late Cenozoic-Recent Pectinidae (Mollusca: Bivalvia) of the Southern Ocean and neighbouring regions. Monographs of Marine Mollusca, 5: i-viii + 1-125 pp, 17pls. Backhuys Publishers, Leiden [p. 51]
|

Psychrochlamys delicatula delicatula (Hutton); H. A. Jonkers, 2003, Late Cenozoic-Recent Pectinidae (Mollusca: Bivalvia) of the Southern Ocean and neighbouring regions, plate 10, figures e-g.
|
|
«REMARKS: The taxonomic position and specific status of this form have been debated for some years in New Zealand since Fleming (1944) pointed out its paleoclimatic significance, and (Fleming 1944, p. 211) suggested (because Ihering (1907) included C. patagonica in his subgenus Zygochlamys) that Zygochlamys lhering, 1907 might be a suitable subgeneric position for it. Morra (1985) specifically excluded the South American post-Oligocene species from Zygochlamys, and Del Rio (1992, p. 25-30) discussed the relationships and evolution of species related to C. actinodes (Sowerby), which she followed Morra (1985) in placing in a group "Chlamys" distinct from Zygochlamys. Waller (1991, p. 28-29) gave firm phylogenetic reasons for placing all these species, including Recent ones, in the genus Zygochlamys, and provided characters separating Z. delicatula from Z. patagonica at the species level. As noted above, the figures of left valves in Morra (1985) and Del Rio (1992) leave no doubt that all the Argentinian Eocene to Miocene species are congeneric with "Chlamys" patagonica, so Zygochlamys is accepted as the correct generic name for what is now a cold-water, circum-subantarctic group of large Chlamydinae. Waller's opinion on the specific status of Z. delicatula is also accepted here; any closer analysis of the relationships of the living forms requires electrophoretic comparison, beyond the scope of the present project.
An important character for recognising this genus is its left valve preradial dissoconch microsculpnue of simple Camptonectes antimarginal ridglets (Figure 3b,c), without obvious commarginal ridges. Others are the lack of shagreen microsculphue at all growth stages (in Miocene and younger taxa; see above), and the regular, fine commarginal ridges and scales of secondary and tertiary costellae and their interspaces being finely fluted by antimarginal ridglets (Fig. 3d; Waloszek 1984, pl. 6, fig. 5).» BEU, A. G. 1995. Pliocene Limestones and their scallops. Lithostratigraphy, pectinid biostratigraphy, and paleogeography of eastern North Island late Neogene limestone. Institute of Geological & Nuclear Sciences Monograph 10. Institute of Geological and Nuclear Sciences, Ltd., Lower Hutt, New Zealand, 243 pp. [p. 16]
|

Zygochlamys delicatula (Hutton); A. G. Beu, 1995, Pliocene Limestones and their scallops, figures 3a-d.
|
|
«ln New Zealand, C. patagonica delicatula lives abundantly today around Macquarie, Campbell, Auckland, Bounty, and Antipodes lslands (Dell, 1964, p. 180) and is commonly dredged alive on the outer continental shelf and in the upper bathyal zone (in depths of about 80 to 500 m) around southern South lsland, as far north as Otago Peninsula. Only occasional non-breeding individuals have been found alive as far north as Cook Strait (Pantin, 1963). As a fossil, C. patagonica delicatula has become important for recognising glacial periods in marine Late Pliocene and Early Pleistocene rocks well to the north of its present distribution, when favourable palaeogeographic conditions and severe cooling of the sea allowed the eastern South lsland current to carry planktonic larvae of subantarctic marine animals as far north as central North lsland (Fleming, 1944, 1953; Beu et al., 1977, 1981; Hornibrook, 1980 p. 118; 1981, p. 288; Edwards et al., 1981).
As a fossil in central New Zealand, Chlamys patagonica delicatula is recorded earliest in the Late Pliocene (late Mangapanian or possibly earliest Nukumaruan) Bull Creek Limestone (Vella and Briggs, 1971, p. 270) in southern Ruakokopatuna Valley, southern Wairarapa. Thereafter it is abundant in a large number of early to middle Nukumaruan (latest Pliocene and Early Pleistocene) localities in North Canterbury, southern Marlborough, Wairarapa, southern Hawkes Bay, the Ruahine Range, and the Hautawa Shellbed in Wanganui Basin (Fleming, 1944; 1953, p. 131; Beu et al., 1977,1981). lt probably occurs in late Nukumaruan rocks in North Canterbury and southern Marlborough, as it is recorded from all demonstrably Nukumaruan rocks in these areas. The synonymy above, records it newly from (probably early) Nukumaruan rocks of Chatham lsland. lt is also recorded from a sinqle Castlecliffian locality (Pecten modestus kupei Zone; Beu, 1977, p.202) in North Canterbury. lts occurrence in 20 000-year-old concretions in Cook Strait was described above; Fleming (1943, p. 193, footnote) and Powell (1950, p. 75) earlier suggested that Cook Strait records were based on fossils from submarine outcrops (which, however, has proved to apply to only some of the Cook Strait records). The long synonymy above attests to the significance that C. patagonica delicatula has assumed in the interpretation of New Zealand Plio-Pleistocene geology. A number of inclusions in the synonymy and deletions from it call for comment. Although some workers as recent as Powell (1979, p. 379) have continued to include Pecten diffluxus Hutton (1873, p. 31) in the synonymy of C. patagonica delicatula, Thomson (1919, p. 282) and Boreham (1961, p. 659, pl. 45, figs 13) long ago showed this to be incorrect; Sectipecten diffluxus (Hutton) is the earliest member of the Sectipecten lineage, of Waiauan (late Middle, or early Late Miocene) age. Chlamys titirangiensis Marwick, 1928 is included in the synonymy on the basis of the very close resemblance of the holotype and two paratypes (NZGS, TM4207) to three abraded valves of C. patagonica delicatula collected recently by Dr P. A. Maxwell from the Titirangi Sand in a large quarry near the new airport on Karewa Peninsula, Chatham lsland. Marwick's types consist of a large valve agreeing in shape with C. patagonica delicatula but with all sculpture eroded off, and two fresh, coarsely sculptured, smaller valves (the larger the holotype); all three are from the Titirangi Sand (Nukumaruan) at Titirangi Point, north shore of Karewa Peninsula where, however, the species is rare; no specimens were found during recent recollections. lt appears that C. patagonica delicatula lived off-shore at the Chatham lslands during the time of deposition of Titirangi Sand, a few eroded valves being contributed to the shallow-water deposition site of the unit. Specimens from Campbell lsland identified by Fleming (1950, p. 49) as 'Chlamys aff. delicatula' have been reexamined, as these specimens would be the earliest record of the species group outside South America if the identification were confirmed. The specimens are from the Shoal Point Formation which also contains calcareous nannofossils. These include Reticulofenestra pseudoumbilica but not Cyclicargolithus floridanus, indicating a probable lower age limit of upper Lillburnian (Middle Miocene - 14 Ma) and a definite upper age limit of mid Opoitian (Early Pliocene - 3.5 Ma) (A.R. Edwards, NZGS, pers. comm.). Most overlying volcanic rocks range in age from 6.0 to 7.5 Ma (Late Miocene) but a few flows from near the base of the sequence yielded ages of 8.5 Ma and 11 Ma (Late and Middle Miocene) (Adams et al., 1979). An upper Lillburnian or Waiauan age (late Middle, to early Late Miocene) is indicated for the Shoal Point Formation. Although all Chlamys specimens in the Shoat Point collection are extremely poorly preserved, they can be seen to reach a height of only about 30 mm, have very weakly inflated discs, and show the internal impressions of numerous, fine, narrow radial ribs; they demonstrate no criteria to differentiate them from the group of species related to the abundant New Zealand late Cenozoic and living species C. gemmulata (Reeve). The record by Kingma (1958, p. 495) consists of large specimens of Chlamys gemmulata from the Waipipian (mid Pliocene) lower limestone of the Te Waka Range, south of the Napier-Taupo Road, Hawkes Bay (Beu et al., 1981, p. 127). The record by Boreham (in Hay et al., 1970, table 4, p. 44) is discussed more fully below; it refers to an unnamed species intermediate between C. seymouri Marwick and C. patagonica, occurring in the Whenuataru Tuff (mid to Late Pliocene) of Pitt lsland, Chatham lslands, and has been deleted from the synonymy. The synonymy list is therefore considered to be reasonably exhaustive for major citations, but a large number of mere faunal listings have not been included.» BEU, A. G. 1985. Pleistocene Chlamys patagonica delicatula (Bivalvia: Pectinidae) off southeastern Tasmania, and history of its species group in the Southern Ocean. In: Lindsay, J. M. (ed. ), Stratigraphy, paleontology, malacology. Papers in honour of DrNell Ludbrook. Department of Mines and Energy, South Australia. Special Publication. 5: 1-11, pls. 1, 2. [p. 6, 7]
|


Chlamys patagonica delicatula (Hutton); A. G. Beu, 1985, Pleistocene Chlamys patagonica delicatula off southeastern Tasmania, and history of its species group in the Southern Ocean, plate 1, figures 5-9; plate 2, figures 1-5 (figures 2-5: paratypes of Chlamys instar Iredale)
|
|
«The extant species that is most closely related to Zygochlamys patagonica is Z. delicatula (Hutton, 1873) (PI. 2, Figs. 15, 16), which lives in sub antarctic New Zealand waters around Macquarie, Campbell, Auckland, Bounty, and Antipodes Islands (Pantin, 1963; Dell, 1964; Beu, 1985). Beu (1985) also reported that this species "is commonly dredged alive on the outer continental shelf and in the upper bathyal zone (in depths of about 80 to 500 m) around southern South Island (New Ze aland), as far north as Otago Peninsula." Although the species ranges as far north as the Cook Strait, at the north end of South Island, abundance declines very rapidly north of the Otago Peninsula (Panlin, 1963).
The New Zealand and South American taxa differ from one another in a number of fcatures: (1) Z. delicatula has a ribbing pattern that is more regular, with each major rib in the center of the disk of each valve bearing a row of scales or a scale-topped second ary riblet on each of its flanks; in Z. patagonica, the ribbing pattern is more irregular, and such tripartite ribs are far less common. (2) The byssal notch of the right valve of Z. patagonica is shallower in mature specimens than is that of Z. delicatula, and a corresponding difference occurs on the left valve, with the byssal sinus of Z. patagonica being non-existent, the free margin of its left anterior auricle tending to form a 90º or obtuse angle with the hinge line. (3) Z. patagonica is generally less brilliantly colored, having a left valve that is gen erally a dull brick red in color, in contrast to the left valve of Z. delicatula, which is commonly a brilliant hue of yellow or orange. (4) Z. delicatula commonly has an early growth stage having commarginal lamellae in the rib interspaces and high narrow ribs, with scales beginning abruptly but at different times on each rib; in Z. patagonica this early lamellate stage is either absent or poorly demarcated, with lamellae tending to appear in interspaces at irregular intervals. Both taxa exhibit a remarkable tendency for commarginal lamellae or scale s to be fluted by antimarginal ridglets (see Waloszek, 1984, pI. 6, fig. 5), and both also show exactly the same degree of foliated calcite reentry on the shell interior. This reentry extends below the level of the adductor scar but does not quite occupy the entire space within the pallial line. Beu (1985) maintained that the differences in ribbing, shape, and color that separate the New Zealand and South American taxa are intergradational and concluded that these taxa should be treated as geographical subspecies, Chlamys patagonica patagonica and C. patagonica delicatula. He maintained that the intergradation was evidence of gene flow through the agency of planktonic larval dispersal via the eastward flowing Antarctic Circumpolar Current. On the basis of the ages of related fossil species, Beu also concluded that the Z. patagonica species complex originated in South America in the Miocene, became distributed around the Southern Ocean, and finally became adapted to cold conditions in the late Pliocene. He rejected the notion that Pecten geminatus Sowerby, 1846, a late Eocene or early Oligocene fossil from Argentina (figured by Zinsmeister, 1981) is ancestral to the Z. patagonica complex. Because this fossil species is the type species of Zygochlamys Ihering, 1907, Beu therefore rejected the applicat ion of the name Zygochlamys to this group, choosing instead to place the group in the genus Chlamys, sensu lato.» WALLER, T. R. 1991. Evolutionary relationships among commercial scallops (Mollusca: Bivalvia: Pectinidae). In: Shumway S. E. (ed.), Scallops: biology, ecology and aquaculture. Developments in Aquaculture and Fisheries Science, Elsevier, Amsterdam, 21: 1-73, pls. 1-8. [pp. 28, 29]
|

Zygochlamys delicatula; T. R. Waller, 1991, Evolutionary relationships among commercial scallops, plate 2, figures 15, 16.
|
|
«REMARKS: The New Zealand Chlamys delicatula, including the Recent C. subantarctica Hedley and C. campbellica Odhner (Powell, 1950, p. 76), is thought to be related to a group of large, free-living Chlamys, including the Patagonian C. geminata (Sowerby), type species of Zygochlamys, which have had a circumpolar, predominantly antarctic and subantarctic, distribution during late Tertiary to Recent times, (Fleming, 1944, 1957; Powell, 1960, 1955). The earliest known representative of the Zygochlamys group in New Zealand is Chlamys (Z.) seymouri Marwick from the Opoitian of the Chatham Islands.»
BOREHAM, A. U. E. 1965. A revision of F. W. Hutton's pelecypod species described in the Catalogue of Tertiary Mollusca and Echinodermata. New Zealand Geological Survey Paleontological Bulletin, 37: 1-125, pls. 1-20. [p. 23]
|

Chlamys (Zygochlamys) delicatula (F. W. Hutton, 1873); A. U. E. Boreham, 1965, A revision of F. W. Hutton's pelecypod species described in the Catalogue of Tertiary Mollusca and Echinodermata, plate 5, figures 8-10.
|
   |