Crassadoma gigantea (Gray, 1825)
GRAY, J. E. 1825. A list and description of some species of shells not taken notice of by Lamarck. The Annals of Philosophy [New Series], 9: 134-140. [p. 139]
1825 Lima gigantea Gray, 1825
1834 Hinnita poulsoni Conrad, 1834
1857 Hinnites crassa Conrad, 1857
1928 Pecten (Chlamys) multirugosus Gale, 1928
1928 Pecten (Chlamys) multirugosus var. crassiplicatus Gale, 1928 [nomen novum pro Hinnites crassa Conrad, 1857]
1834 Hinnita poulsoni Conrad, 1834
1857 Hinnites crassa Conrad, 1857
1928 Pecten (Chlamys) multirugosus Gale, 1928
1928 Pecten (Chlamys) multirugosus var. crassiplicatus Gale, 1928 [nomen novum pro Hinnites crassa Conrad, 1857]

Pecten (Hinnites) giganteus Gray, H.C. Küster & W. Kobelt, 1888, Die Gattungen Spondylus und Pectens, plate 66, figures 1-3.
|
«LIMA gigantea. Testa crassa, ponderosa, subauriculata. albido-rosea, irregulariter radiata costata striata; intus alba, rufo maculata. lat. 15.4, long. 18.4 unc.»
JOHN EDWARD GRAY, 1825
|
















|
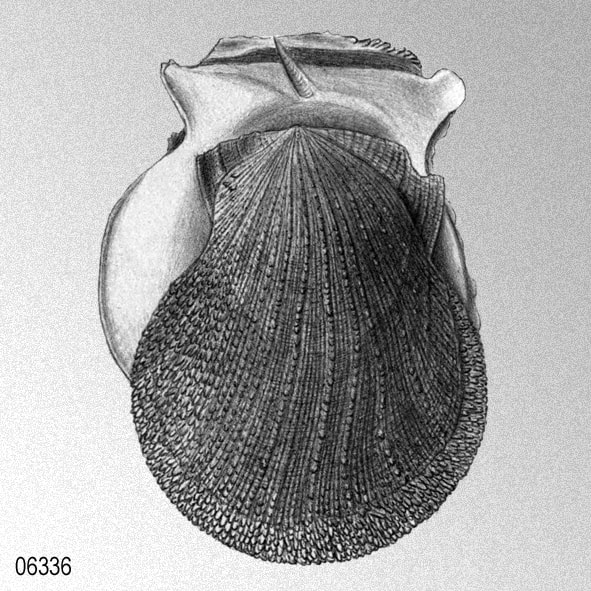
«Crassadoma gigantea (J. E. Gray, 1825). Giant rockscallop.
Lima gigantea J. E. Gray 1825; Hinnita poulsoni Conrad, 1834; Hinnites crassa Conrad 1857; Pecten (Chlamys) multirugosus Gale, 1928; P. (Chlamys) multirugosus crassiplicatus Gale 1928. Type locality: Juan de Fuca Strait. Distribution: Montague Strait, Prince William Sound, Alaska, south to Bahia Magdalena, Baja California Sur, and on Isla Guadalupe, Baja California; intertidal zone to 80 m.» DRUMM, D. T., K. P. MASLENIKOV, R. V. SYOC, J. W. ORR, R. R. LAUTH, D. E. STEVENSON & T. W. PIETSCH. 2016. An annotated checklist of the marine macroinvertebrates of Alaska. NOAA Professional Paper NMFS, 19: 1-289. [p. 142]
|
|
«The genus Crassadoma was created by Bernard (1986) to accommodate Hinnites giganteus (the former name of Cr. gigantea), a species that shows a cementing life habit, on the grounds that cementing was obligatory in this species of Hinnites, while it was facultative in other Hinnites. However, Harper (1991, cited in Waller 1993) pointed out that the cementing style was the same in giganteus as in the remaining Hinnites. Waller (1993) extended the genus Crassadoma to include other cemented and non-cemented Hinnites species, and created the tribe Crassadomini, which included the genus Caribachlamys as well.
Our analysis indicates that the American Cr. gigantea is closer to Chlamys islandica than to the other member of the Crassadoma genus included in this study (Cr. multistriata, from the eastern Atlantic and western Indian Ocean). Cr. multistriata, in turn, has Mimachlamys varia as sister species. These results have implications regarding the origin of C. gigantea. Grant and Gale (1931) and Waller (1991) assumed that this species originated in the eastern Pacific from a member of the Chlamys group, presumably C. hastata. Waller (1993) later suggested that an origin from an Atlantic species would be more likely, given the highly derived traits of C. gigantea in comparison with other Chlamydinae and the absence of Crassadoma in the Indo-west Pacific. Our results clearly support that C. gigantea originated from the C. islandica group of species, to which C. hastata belongs (Waller 1991). Our results also support the view that the cementing habit appeared independently in several lineages (Waller 1991, 1993). However, these lineages were not different waves of Crassadoma moving from the eastern Atlantic to the western Atlantic and then to eastern Pacific, as suggested by Waller (1993), but different Chlamydinid lineages. Finally, our results agree with the view of Hinnites as a polyphyletic genus (Waller 1991, and references therein), and indicate that Crassadoma is also polyphyletic.» SAAVEDRA, J. & J. B. PEÑA. 2006. Philogenetics of American scallops (Bivalvia: Pectinidae) based on partial 16S ribosomal RNAgene sequences. Marine Biology, 150: 111-119. [p. 114, 115]
|

Crassadoma gigantea (Gray, 1825); B. K. Raines & G. T. Poppe, 2006, A Conchological Iconography. The Family Pectinidae, plate 206, figures 1-4.
|
|
«Comparison.— Crassadoma gigantea differs from its cemented congener of the eastern Atlantic, C. pusio (see below), in being of much greater maximum size and in having more prominently fasciculated ribs on its right valve and more distinctly ordered ribs on its left (compare Figs. 6a, b with Figs. 6k, 1). The vivid purple color present on the hinge plate of C. gigantea, particularly on the left valve, is absent or faint in C. pusio. Although it is generally assumed that C. gigantea is thicker shelled than is C. pusio, it is doubtful that there is any significant difference when size is taken into account. Both C. gigantea and C. pusio have extensive development of foliated calcite inside the pallial line, and in both species the crossed lamellar aragonite that is present in this region in early ontogeny is covered over by foliated calcite earlier on the left valve interior than on the right, a common phenomenon among pectinids. C. gigantea is more derived than C. pusio in the sense that the foliated calcite transgression begins earlier in ontogeny, appearing on the umbonal interior of right valves as small as 11 mm in shell height and on left valves as small as 7 mm. In C. pusio it has been observed in right valves no smaller than 17 mm and on left valves no smaller than 13 mm. In C. multistriata the plesiomorphic condition persists; there is no foliated calcite transgression over the umbonal interiors at any stage of ontogeny.
Adegoke (1969: 103) described a new species of "Hinnites", Hinnites benedicti Adegoke, 1969, from the Late Miocene Santa Margarita Formation of California. Although this species was found in the same beds as H. multirugosus crassiplicatus (Gale, 1928) [= Crassadoma gigantea], Adegoke regarded it as distinct because of its larger Chlamys stage and more even ribbing. My own examination of Adegoke's types at UCW confirmed this distinctness but did not confirm that "Hinnites" benedicti has a cementing habit. The maximum size of the Chlamys stage (34 mm) given by Adegoke is exceeded by the height of the holotype (46.8 mm) and two of the paratypes, none of which show any evidence of cementation. The holotype, a right valve, would be expected to show an abrupt sculptural change and xenomorphic growth if cemented. Instead, the degree of shell irregularity is comparable to that of extant Laevichlamys irregularis of the Indo-Pacific, a species with a byssate, nestling living habit. The style of ribbing of H. benedicti and the apparent lack of commarginal lirae suggest that it is not a Crassadoma, but its generic assignment awaits examination of better preserved material. Most likely it is a Chlamys, s.s. on the basis of its style of ribbing introduction. Living habits.— From just below low tide level to 80m (Hertlein, 1972: 212; Grau, 1959: 137; Bernard, 1983: 25); attached to rocks, coarse gravel, and other hard objects. Specimens in northern waters tend to live closer to shore and at shallower depths than those in southern waters (Hertlein, 1972: 212). Geographic range.— Aleutian Islands, Alaska, to Bahia Magdalena, Baja California, Mexico; offshore in Santa Barbara Islands, California, Guadalupe Island, Mexico (Grau, 1959: 137), and Islas de Revillagigedo, Mexico (reported herein, CAS 60266). Stratigraphic range.— Early Miocene?, Middle Miocene to present. The presence of Crassadoma gigantea in the Middle Miocene through Pleistocene of California is apparently well established (Moore, 1984: 66). Lower Miocene records, however, are few in number and could require reexamination. The only Lower Miocene record listed by Moore (1984: 66) is from the Painted Rock Member of the Vaqueros Formation. Arnold (1906: 94) also listed a single occurrence, "associated with Turritella hoffmanni" in rocks that he referred to as Lower Miocene. Smith (1991a: 35), in reviewing the changing concepts of the age of the "Vaqueros Stage", pointed out that some records in faunal lists need redetermination and further collecting to determine stratigraphic position. The earliest reliable records in California appear to be from the "Margaritan Stage", which Smith (1991a, figs. 12, 14) shows as ranging from planktic foraminiferal zone N12 into lower N16 (Langhian into lower Tortonian stages of Europe) and as correlating with the Shoal River and Yellow River Formations of Florida. Discussion.— Previous workers (e.g. Grant and Gale, 1931: 161; Waller, 1991: 23) have assumed that Crassadoma gigantea evolved from a member of the Chlamys group, e.g. Chlamys hastata, within the eastern Pacific. For reasons given above, however, this is unlikely. C. hastata and its congeners in the eastern and northern Pacific have features that place them squarely within the tribe Chlamydini as herein defined, whereas C. gigantea has a microsculptural pattern shared with the other species placed here in the tribe Crassadomini. Specifically, in the early ontogeny of C. gigantea the plesiomorphic pattern of continuous sweeping antimarginal striae is absent and commarginal lirae are prominent in rib interspaces (Fig. 5f). Crassadoma gigantea has a ribbing pattern that is more derived in comparison to the patterns of its eastern Atlantic congeners, and its large size and purple hinge also are derived in comparison to the eastern Atlantic species and to outgroup Chlamydinae. Given the absence of Crassadoma in the Indo-west Pacific, it is highly likely that C. gigantea evolved from species that were present in the Atlantic. There remains the problem, however, of where the fossils are which should corroborate this ancestry. If it is true that C. gigantea was already present in the eastern Pacific by the middle Miocene, it is likely that an ancestor was present in the western Atlantic and that it dispersed through seaways to the Pacific at this time or earlier. If the ancestral species was eurytopic with a preference for warm temperate but not strictly tropical temperatures, it is possible that it avoided the shallow water habitats that predominate in the early Miocene stratigraphic sections thus far known in the Caribbean. The absence of these fossils thus could be the result of a facies sampling bias in the region in which they are most likely to be found. The most likely interval of the middle Miocene for speciation to have occurred may have been during the time of initial emergence of the Isthmus of Panamá. This uplift cut off the flow of intermediate water between the Atlantic and Pacific and intensified the southward flow of the cool California Current along the Mexican coast, creating a thermal barrier to the west-to-east dispersal of shallow water taxa (Duque-Caro, 1990).» WALLER, T. R. 1993. The evolution of Chlamys (Mollusca: Bivalvia: Pectinidae) in the tropical western Atlantic and eastern Pacific. American Malacological Bulletin, 10 (2): 195-249. [p. 211, 212]
|
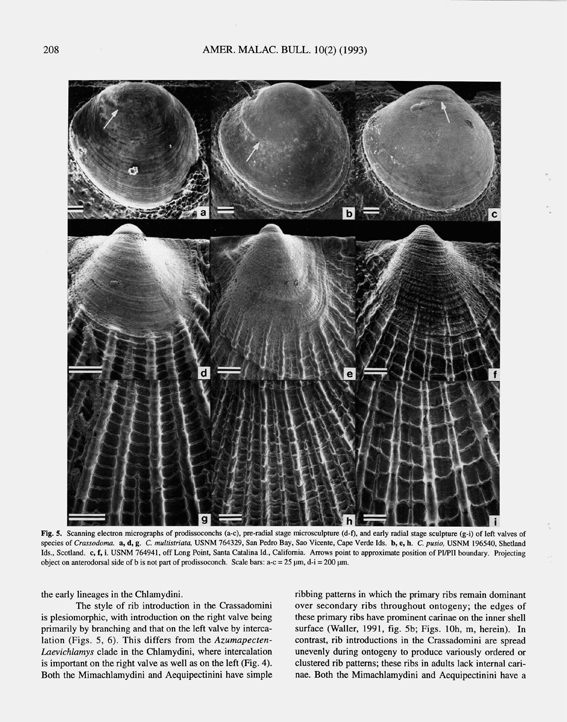

Crassadoma gigantea (Gray, 1825); T. R. Waller, 1993, The evolution of Chlamys, figures 5c, 5f, 5i; figures 6a, 6b.
|
|
«THE COMMON ROCK SCALLOP, or "purple-hinged scallop," of the Pacific Coast appears in recent literature under two names: Hinnites giganteus (Gray, 1825) and Hinnites multirugosus (Gale, 1928). GALE (1928: 92) introduced the latter because he believed that Gray's name and other possible substitutes were homonyms and therefore unavailable. Other authors (ADAM, 1960; HERTLEIN & GRANT,1972) have given reasons for preferring use of the oldername, H. giganteus. The valid name of this taxon depends on two related factors: (a) the availability, or not, of Gray's name, and (b) the nature and validity of Gale's rejection of it— both points to be evaluated in light of the ICZN rules governing such cases.
From a review of the relevant literature, we have come to the following conclusions: I. GALE's (1928) stated reasons for proposing "Pecten (Chlamys) multirugosus" — a new taxon, not a simple renaming— and his remark that Lima gigantea Gray was "stillborn" are erroneous. Lima gigantea GRAY (1825: 139), as the species was first named, is the earliest use of this combination of generic and specific names. It is not a primary homonym of Plagiostoma giganteum J. Sowerby, 1814, whether or not Lima Bruguière, 1797, and Plagiostoma J. Sowerby, 181 4, are considered synonymous genera of the Limidae. GRAU's (1959) claim that Plagiostoma was proposed as a subgenus of Lima is incorrect. Moreover, GRAY (1826) removed his gigantea from Lima and placed it in "Hinnita" (an invalid emendation of Hinnites Defrance, 1821), and the species has not been reallocated to Lima since that time. We have located no citation of Sowerby's giganteum in the genus Lima prior to those by DESHAYES (1831, 1832), although the possible synonymy of Lima and Plagiostoma was being debated as early as 1823 (G. B. SOWERBY, 1823) . Modem workers consider the two genera separable. As ADAM (1960) noted, Gray's and Sowerby's names were not secondary homonyms at the time of GALE's (1928) publication. It is evident that Gale himself did not believe the two species to be congeneric, since he cited Sowerby's giganteum as a Lima but described multirugosus as a species of Pecten. Under ICZN rules [Art. 59(b)(2)], Gray's name required no replacement then and requires none today. 2. JAY (1835) and many subsequent authors placed Gray's gigantea in the genus Pecten Müller, 1776. While several other pectinid taxa share the specific epithet "giganteus," the earliest of these is Pecten giganteus Münster in Goldfuss, 1833. The gigantea of GRAY (1825) is therefore not a junior secondary homonym in Pecten. 3. GALE (1928) specifically stated that his Pecten (Chlamys) multirugosus was proposed as a new species, and not the simple renaming of a homonym: "Pecten multirugosus is virtually a new name for the common Pliocene to Recent West Coast species formerly known as Pecten (Hinnites) giganteus (Gray); but in order to avoid any questions about the location or identity of the original types, the species is described as new and a new type is cited" (GALE, 1928: 92; emphasis supplied). In contrast, "Pecten (Chlamys) multirugosus var. crassiplicatus," a renaming of the homonymous Hinnites crassa Conrad, 1857, by Gale in the same paper, was unequivocally proposed as a replacement name. 4. In summary, GRAY's (1825) name gigantea was available when proposed and has remained available throughout its nomenclatural history. GALE's (1928) rejection of it was invalid because he did not contend that the two species-group taxa, "Lima" gigantea Gray and Plagiostoma giganteum Sowerby, were congeneric. Pecten (Chlamys) multirugosus Gale, 1928, is therefore a junior synonym of Hinnites giganteus (Gray, 1825), and the latter is the valid name for the Pacific Coast rock scallop.» ROTH, B. & E. V. COAN. 1978. Nomenclatural notes on Hinnites giganteus (Gray). The Veliger, 20 (3): 297-298. [p. 297]
|
Hinnites giganteus Gray; L. G. Hertlein & U. S. Grant IV, 1972, The Geology and Paleontology of the marine Pliocene of San Diego, California, plate 41, figure 16.
|
|
«Hinnites crassa Conrad (470), described from beds of late Miocene age in California was renamed crassiplicatus by Gale (471) as a variety of the Recent form multirugosus. However, the name of Conrad's species is not preoccupied by Pecten crassus Risso, 1826, and no replacement is necessary. Gale believed the Miocene form to be smaller, usually with the Chlamys stage lasting longer, with less irregular growth and with fewer more highly differentiated coarse, rugose radial ridges. The taxonomic value of the name applied to this form is open to question. It may be a subspecies of the Recent form but Arnold (472) considered it to be identical with the Recent species and we are inclined to that view. Woodring and Stewart (473) reported "Hinnites cf. H. crassa Conrad" from the Etchegoin Formation, late Pliocene, in the San Joaquin Valley. Hinnites benedicti Adegoke (474), from the Santa Margarita Formation of late Miocene age, was described from the same region.
Dollfus and Dautzenberg (475) mentioned that a series of specimens of Hinnites crispa from late Tertiary beds in Europe reveals variations analogous to those of H. giganteus.» HERTLEIN, L. G. & U. S. GRANT IV. 1972. The Geology and Paleontology of the marine Pliocene of San Diego, California (Paleontology: Pelecypoda). Memoirs of the San Diego Society of Natural History, Memoir 2, Part 2b. pp. 143-409, pls. 27-57. [p. 212]
|

Hinnites cf. H. crassa Conrad; W. P. Woodring, R. Steward & R. W. Richards, 1972, Geology of The Kettleman Hills oil field California, plate 31, figures 3, 4, 8.
|
|
«Pecten (Chlamys) multirugosus var. crassiplicatus GALE, 1928, p. 93.
Sculpture consists of more than 17 primary ribs with well-developed secondary ribs occurring singly or in pairs between the primary ribs. In a few interspaces one or two tertiary ribs are also present.
The specimens resemble H. crassiplicatus more than they resemble the Recent specimens of H. multirugosus, which bear as many as five secondary ribs between pairs of primary ribs. Until it is demonstrated that the Miocene to Recent Hinnites of the Pacific Coast may be part of a single variable group, it is useful to point out the sculptural differences of Miocene specimens by use of the varietal name.» STANTON JR., R. J. 1966. Megafauna of the upper Miocene Castaic Formation, Los Angeles County, California. Journal of Paleontology, 40 (1): 21-40, figs. 1-2, pls. 5-7. [p. 27, 28]
|

Hinnites multirugosus var. crassiplicatus (Gale),; R. J. Stanton Jr., 1966, Megafauna of the upper Miocene Castaic Formation, Los Angeles County, California, plate 5, figure 6.
|
      |


