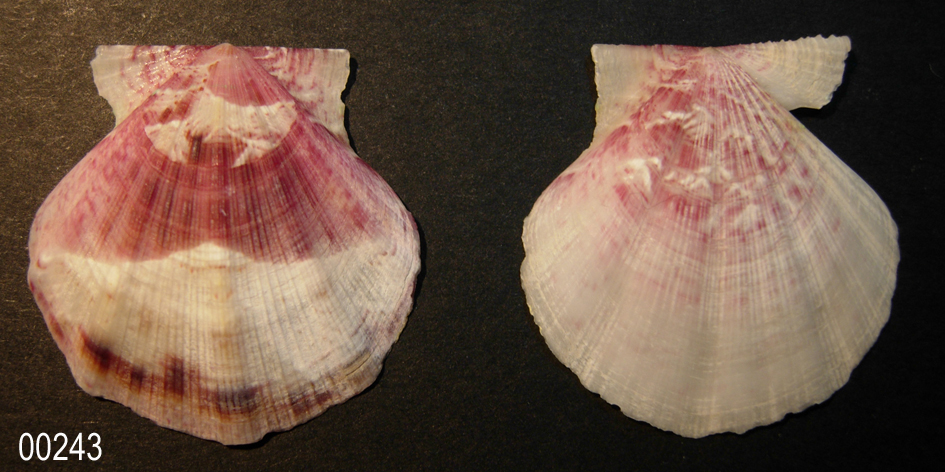
Flexopecten glaber proteus (Solander in Dillwyn, 1817)
DILLWYN, L. W. 1817. A descriptive catalogue of Recent shells, arranged according to the Linnaean method; with particular attention to the synonymy. Vol. I. 1-580 pp. Printed for John and Arthur Arch, Cornhill. London. [p. 265]
1817 Ostrea proteus Solander in Dillwyn, 1817

Pecten discors seu disconveniens; J. H. Chemnitz, 1795, Neues Systematisches Conchylien-Cabinet, Band 11, plate 207, figure 2042.
|
«PROTEUS. 41. Shell sub-orbicular, with about five broad convex plaits, and numerous longitudinal striae; auricles nearly equal.
Ostrea Proteus. Solander's MSS. Pecten discors, seu disconveniens. Chemnitz, xi. p. 264. t. 207. f. 2042. Inhabits the Mediterranean. Solander. Shell about an inch, or sometimes an inch and a half long, and the breadth exceeds the length about two lines; the colour of the upper valve is reddish or pale brown, sometimes marked with darker transverse zones, and sometimes slightly clouded with white; the lower valve is generally white.» LEWIS WESTON DILLWYN, 1817
|




|
«Chlamys proteus or F. proteus Dillwyn has been considered since long, as a species of the bivalve family Pectinidae (Cossignani et al., 1992; Delamotte & Vardala–Theodorou, 1994; Doneddu & Trainito, 2005; Repetto et al., 2005; Zenetos et al., 2005). The majority of these sources mention that F. proteus is another similar species to F. glaber. The main systematic characteristic for the recognition of the two pectinids, is the difference in the number of major and dominated ribs which are 9-12 in F. glaber, but only five in F. proteus (Fig. 2). On the other hand, according to Poppe & Goto (1993), CLEMAM & ERMS, F. proteus is accepted as a subspecies or a form of F. glaber. Also, Raines & Poppe (2006) underline that F. proteus “is treated only as a synonyme” of F. glaber.

Many biologists have argued that the future of descriptive taxonomy will depend on successfully embracing new techniques. Many ideas have been proposed and much progress has been achieved by using molecular data that provide a complementary approach to discriminate species separated by subtle morphological characters (Knowlton, 1993; Avise, 1994; Chan & Chu, 1996; Sarver et al., 1998; Mathews et al., 2002; Goetze, 2003). The invertebrate mtDNA 16S rDNA gene has been relatively well studied due to availability of universal primers (Kocher et al., 1989; Palumbi, 1996) and was used to resolve taxonomic problems in the family Mytilidae (Rawson & Hilbish, 1995), Veneridae (Canapa et al., 1996) and Pectinidae (Canapa et al., 2000a; Saavedra & Pena, 2004; Pujolar et al., 2010).
Taking into account all the existed considerations and the uncertain systematic status of these two common Flexopecten “forms”, another aim of our work was an attempt to solve the identification problem of the two Aegean taxa, using the sequence analysis of the mtDNA 16S rDNA gene. A parallel and similar study (Pujolar et al., 2010), was also made for the Adriatic populations of the Flexopecten complex.
[...]
We developed a DNA methodology based on PCR amplification and sequencing analysis of the mitochondrial 16S rDNA gene, in order to discriminate two different taxa of the family Pectinidae - Flexopecten glaber and Flexopecten proteus - and for a population study of both taxa. Fifty polymorphic nucleotide sites and fifty one different haplotypes were revealed in total. The majority of the genetic variation (96.98%) was distributed within populations. The levels of haplotype and nucleotide diversity were relatively high for both taxa, probably due to the large effective population sizes and the good population status. No species specific position was found and the two highly frequent haplotypes are shared between taxa, suggesting possible hybridization between them. Genetic distance between F. glaber and F. proteus was D = 0.005 and no genetic differentiation was revealed among taxa, when pooling together samples from different localities. Our results show that F. glaber and F. proteus are conspecific, which is in agreement with the latest classification.»
IMSIRIDOU, A., N. KARAISKOU, E. AGGELIDOU, V. KATSARES & S. GALINOU-MITSOUDI. 2012. Mitochondrial DNA variation as a tool for systematic status clarification of commercial species - The case of two high commercial Flexopecten forms in the Aegean Sea. In Zainal Abidin Muchlisin (Ed.): Aquaculture: 109-126. Published by InTech. Rijeka, Croatia, 2012. [p. 111, 122]
|
|
«Description. — Le galbe est un peu triangulaire, les bords latéraux presque rectilignes, descendant au dessous de la mi-hauteur, le postérieur est un peu plus long.
L'oreille postérieure a le bord libre, à peine ondulé, tandis que l'antérieure est pourvue d'un sinus assez aigu. A la surface de la valve droite les 4 larges côtes, peu saillantes, sont très supérieures aux intervalles. Elles sont plus ou moins imparfaitement divisées par lin sillon largement arrondi, pen profond, indiqué dès le sommet. En dehors des côtes principales on tronve de chaque côté encore une petite côte souvent bifide. A la valve gauche, on compte 5 côtes principales, dont 3 dominantes; les deux autres, l'antérieure smtout, sont bifides. Alternant avec les côtes principales, se trouvent des côtes secondaires très faibles, à raison d'une au milieu de chacun des très larges intervalles. Vers le sommet, côtes principales et côtes intercalaires sont peu différentes; on compte donc de 10 à 11 côtes à section passablement carénée; par la suite la prédominance de 5 d'entre elles saffirme. L'ornementation radiale est constituée par des filets, plus ou moins marqués, suivant les individus. La très fine ornementation concentrique est identique à celle de Ch. glabra. Rapports et différences. — C'est de cette dernière que Ch. protea se rapproche le plus. La distinction des formes typiques de l'une et l'antre espèce n'offre pas de difficultés. Par contre dans les var. anisopleura, LOC. de Ch. glabra, et var. praeterita, B. D. D. de Ch. protea, la costulation primaire est bien voisine. Pour les formes se rapportant à Ch. protea, le contour est moins arrondi, les bords latéraux descendent plus bas, l'angle apical est en général plus faible, les côtes de la valve gauche plus inégales, et celles cIe la valve droite plus rapprochées, pIus larges. Répartition géographique et stratigraphique. — Localisée dans la Méditerranée, el surtout répandue dans l'Adriatique actuellement, celle espèce est apparue dans ce domaine sans doule plus tard que Ch. glabra, el y est toujours plus rare. L'une des premières citations est celle du Quaternaire ancien de Casa Campana (Italie méridionale); elle se trouve ensuite dans les couches à Strombus de Tarente. Cependant elle existait déjà dans le Pliocène ancien (var. subpraeterita, SACCO de l'Astesan).» ROGER, J. 1939. Le genre Chlamys dans les formations néogènes de l'Europe. Conclusions générales sur la répartition géographique et stratigraphique des Pectinidés du Tertiare Récent. Mémoires de la Société Géologique de France [Nouvelle série], 17 (2-4):1-294, pls. 1-28. [p. 85, 86]
|

Chlamys protea Sol. var. transiens Gignoux, Quaternaire à Strombus de Tarente (Italie); J. Roger, 1939, Le genre Chlamys dans les formations néogènes de l'Europe, plate 10, figure 10.

Chlamys protea Solander. J. Roger, 1939, Le genre Chlamys dans les formations néogènes de l'Europe, plate 11, figures 3, 3a (actuel, Mediterranée), 4 (Quaternaire ancien de Cutro, Italie).
|
                 |