Caribachlamys pellucens (Linnaeus, 1758)
LINNAEUS, C. 1758. Sistema
Naturae per Regna tria Naturae, secundum classes, ordines, genera, species, cum
characteribus, differentiis, synonymis, locis. Tomus
I. Editio Decima, Reformata, iii, 824 p. Impensis Direct. Laurentii Salvii. Holmiae [Stockholm], 1758. [p. 698]

Peigne semi-auritus; A. J. D. d'Argenville, 1752, L’Histoire naturelle éclaircie dans deux de ses parties principales, la Lithologie et la Conchyliologie dont l’une traite des Pierres et l’autre des Coquillages, plate 27, figure H.
|
«O. testa radiis 9, laevi: fornice squamis cochleari-hemisphaericis.
Argenv. conch. t. 27. f. H. Pecten semiauritus. Habitat in O. australiore.» CAROLUS LINNAEUS, 1758
|




|
«Type locality
“Habitat in O. australiore”. The present species does not occur in the Indo-Pacific. The type locality is designated herein as Florida, U.S.A. Examined material (ZMA) resembles the type material.
Odhner (1953: 6) isolated a specimen with a handwritten Thunberg label ‘‘Mus. Gust.Ad.’’ and identified O. pellucens as a senior synonym of Pecten imbricatus (Gmelin, 1791) on another label (see Wallin, 1993: 87, Ostrea pellucens # 495). This specimen is designated herein as the lectotype (Fig. 10D).»
DIJKSTRA, H. H. 1999. Type specimens of Pectinidae (Mollusca: Bivalvia) described by Linnaeus (1758-1771). Zoological Journal of the Linnean Society, 125: 383-443. [p. 411]
|

Ostrea pellucens Linnaeus, 1758; H. H. Dijkstra, 1999, Type specimens of Pectinidae described by Linnaeus, figure 10D.
|
|
«Type specimens.— The specimen represented by the figure that Gmelin cited, which is in Chemnitz (1784), vol. 7, pl. 69, fig. G, is designated as the lectotype of Ostrea imbricata Gmelin. Many of the specimens illustrated by Chemnitz were from the Spengler Collection housed at the Zoological Museum of Copenhagen (Keen, 1966). It is possible that the illustrated specimen of O. imbricata could be in that collection. I examined specimens labeled as Pecten imbricatus by Lamarck in Paris and Geneva; they all conform to existing concepts of the western Atlantic taxon.
Type locality.— Corrected herein to the Antillean region, western Atlantic. Diagnosis.— Caribachlamys with left valve less convex than right and distinctly flattened; major ribs few in number, commonly 10 on right valve and nine on left, with five ribs on left valve tending to be more prominent and with higher scales than others; secondary riblets variably developed, of much lower height than major ribs; major ribs somewhat trigonal in cross section on right valve, rounded on left; scales on major ribs large, widely spaced, distally concave, commonly forming closed knobs; early commarginal lirae obscured by stronger antimarginal striae and of highly irregular trend; antimarginal striae absent from crests of major ribs. Morphological variation.— Caribachlamys imbricata displays substantial geographic variation, but on the basis of the collections examined this variation appears to be geographically clustered rather than clinal. For purposes of discussion, two intergradational extremes of variation are referred to herein as the Florida morph and the Bermuda morph. Parallel differences between the two are as follows: Florida morph (Figs. 9f, g): (1) In the early ontogeny of the right valve of the Florida morph, secondary costae that are close to or on the flanks of the primary ribs appear by a shell height of 5 mm. By a height of 10 mm, these secondary costae either disappear or are represented by the expanded, rounded rib flanks of the primary ribs; (2) The posterior auricle of the right valve has unevenly distributed costae, the dorsal one or two closer together and much stronger than the others; (3) On the left valve, third-order riblets are usually absent at least in early ontogeny (up to a shell height of 10 mm) and commonly absent throughout ontogeny; (4) On the left valve, there is usually only one riblet present on the anterior side of the anteriormost primary rib in early ontogeny, increasing to two in later growth. These anteriormost riblets lack scales; (5) On the left valve, radial riblets are unevenly distributed on both auricles, the dorsalmost riblets commonly much stronger than the others. Bermuda morph (Figs. 9h-l): (1) On the right valve, secondary riblets are distinct and well separated from the primary ones at a shell height of 5 mm. At a shell height of 10 mm, these secondary costae are still distinct from the major ribs, the major ribs being without expanded flanks; (2) The posterior auricle of the right valve has fairly evenly distributed riblets of even amplitude; (3) On the left valve, third-order riblets are commonly present by a shell height of 10 mm and may be numerous by this stage of ontogeny; (4) On the left valve, there are usually two distinct and fairly strong riblets present on the anterior side of the anteriormost primary rib, commonly increasing to three in later growth. One of these anterior riblets may bear cuspate scales; (5) On the left valve, radial riblets are numerous on both auricles, with only slight or no increase in amplitude toward the dorsal margin. Caribachlamys imbricata is generally lightly pigmented, the darkest parts being the dark reddish or purple maculations that occur in earlier ontogeny between the primary ribs. Solidly colored shells, such as are common for C. sentis, are relatively rare. They have been observed, however, among specimens of the Bermuda morph from the Caribbean coast of Panamá. The valve interiors of both morphs are commonly yellow, sometimes brilliantly so. Comparison.— Caribachlamys imbricata can be distinguished from its congeners by its broad, high-amplitude major ribs. Unlike congeneric species, the scales on these major ribs remain widely spaced throughout ontogeny and commonly form closed knobs. Living habits.— Caribachlamys imbricata lives byssally attached in crevices, beneath coarse rubble and coral heads, and between coral branches on reefs and in both back-reef and fore-reef areas. Depth records for live-collected specimens in museum collections are all shallow, ranging from low subtidal to 20m. Geographic range.— Caribachlamys imbricata appears to have a tropical distribution. It occurs in Bermuda, along the North American mainland only from Miami southward through the Florida Keys, in the Bahamas, and through the Antilles to Curacao and Aruba along the Venezuelan coast. It appears to be absent from the northern Gulf of Mexico, and the only record from the southern Gulf is from Alacran Reef on the north side of the Yucatan Peninsula. Few records are as yet available from the Caribbean coast of Central America with the exception of Belize (offshore cays) and Panamá, which are the best sampled areas. The species is not known from the Atlantic coasts of South America. All specimens of Caribachlamys imbricata from Bermuda examined by me are members of the Bermuda morph, but this morph is also present in Jamaica, Panamá, Colombia, Venezuela, and possibly within samples that also contain the Florida morph in the Bahamas. Stratigraphic range.— Lower? Pleistocene to present. The oldest fossils of Caribachlamys imbricata thus far found are specimens in the Trechmann collection (BMNH) from the Pleistocene Coral Rock of Barbados at "Cane Garden" and "Highgate". These did not appear in the lists published by Trechmann; the species determinations are based on my examination of the specimens in 1979. As reviewed above in the section on C. ornata, Trechmann (1933) considered the Coral Rock to be mainly of early Pleistocene age. I also found fossil Caribachlamys imbricata (USNM(P) 474642) in limestone blocks on Windley Key in the Florida Keys. These blocks presumably came from the Windley Key quarry, which exposes the Key Largo Limestone of late Pleistocene age (Multer, 1969; Stanley, 1966). According to Multer (1969: 109), this limestone represents a back-reef environment and is only about 100,000 years old. Although the specimens are fragmentary, they have ribbing characteristics that suggest that they belong to the Bermuda morph. The only other known occurrences of fossil Caribachlamys imbricata, from Cuba, Haiti, and the Dominican Republic, also appear to be Pleistocene in age (see below under material examined). Discussion.— The two extremes of morphology described above as the Florida morph and the Bermuda morph are of considerable interest in that the former is clearly more derived than the latter when they are compared to outgroup taxa within Caribachlamys, particularly C. mildredae. Among the fossil specimens examined, those from the Windley Key Limestone of Florida (USNM(P) 474642) have ribbing characteristics that suggest that they belong to the Bermuda morph, and this also applies to the specimens from Haiti (USNM(P) 481997 and 481998). The sole Cuban specimen on hand, however, belongs to the Florida morph. A likely explanation for the extant Bermuda populations, which belong exclusively to the Bermuda morph so far as known, is that they represent dispersal to Bermuda early in the evolution of the species at some time during the Pleistocene. Apparently modern oceanographic conditions do not permit continued dispersal of this species to Bermuda, and most members of populations elsewhere have assumed a more derived condition than those in Bermuda. The presence of Bermuda morphs among some extant populations outside of Bermuda suggests that the Bermuda populations should not be referred to as a geographic subspecies.» WALLER, T. R. 1993. The evolution of Chlamys (Mollusca: Bivalvia: Pectinidae) in the tropical western Atlantic and eastern Pacific. American Malacological Bulletin, 10 (2): 195-249, figs. 1-14. [pp. 226-228]
|

Caribachlamys imbricata (Gmelin, 1791); T. R. Waller, 1993, The evolution of Chlamys, figures 9 f-j (f, g, Florida morph; h-l, Bermuda morph).
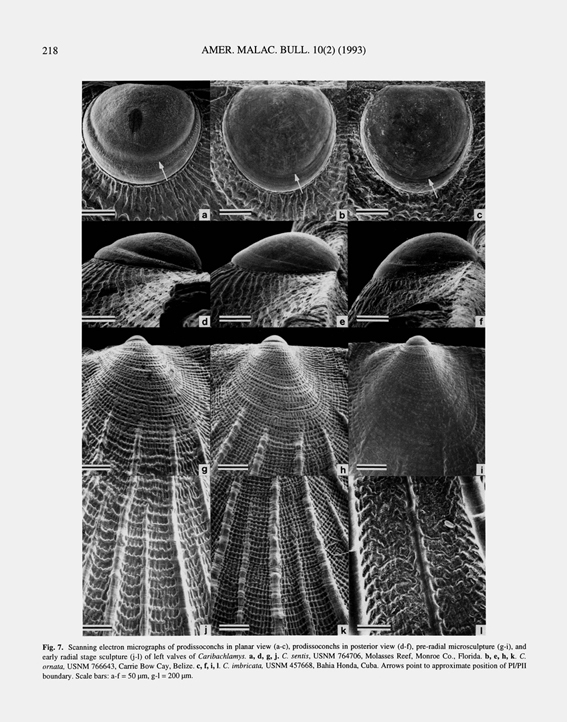
Caribachlamys imbricata (Gmelin, 1791); T. R. Waller, 1993, The evolution of Chlamys, figures 7c, f, i, l.
|
     |