Entolium Meek, 1865
MEEK, F. B. 1865. Description of fossils from the auriferous slates of California. Geological Survey of California, Geology, 1: 477-482, pl. 1 [p. 478]
|
«GENUS AMUSSIUM (Klein), Bolton, 1798.
SUBGENUS ENTOLIUM, Meek. AMUSSIUM AURARIUM, Meek. Plate I, fig. 6, and 6 a. Shell suborbicular, thin, compressed-lenticular, antero-posterior diameter a little greater than that from the hinge to the ventral margin; Sides equally rounded to the regularly rounded ventral border, and converging to the beaks at an angle of about 150°; one ear (the only one known) small, short, fiat, smooth, and obliquely truncated at an obtuse angle of about 125°, to the cardinal margin, which seems to be less than half as long as the breadth of the valves, not defined by even a slight trace of a marginal sinus. Surface of cast, marked with small, obscure, irregular, concentric ridges, separated by wider depressions.
Transverse diameter, 1.86 inches; diameter from ventral margin to beak, 1.67 inches; convexity of the only valve known, 0.18 inch. This species appears to belong to a group of thin lenticular, subequivalve, smooth, or concentrically marked shells, with short, equal, flat, obtusely angular cars, which are not defined in either valve, or on either side, by a distinct marginal byssal sinus. They differ from the typical species of Amussium, in having the lateral margins closed, and the valves without internal radiating costae. This group may be distinguished by the name Entolium, with Pecten demissus, of Phillips (as figured by Quenstedt, in his Der Jura, i, tab. 48, f. 6 and 7), as its type.
Our species differs specifically from Phillips’s in being proportionately broader, straighter on the hinge line, and in having stronger concentric markings.» FIELDING BRADFORD MEEK, 1865
|

Pecten demissus; F. A. Quenstedt, 1856-1858, Der Jura, plate 48, figures 6, 7.
|
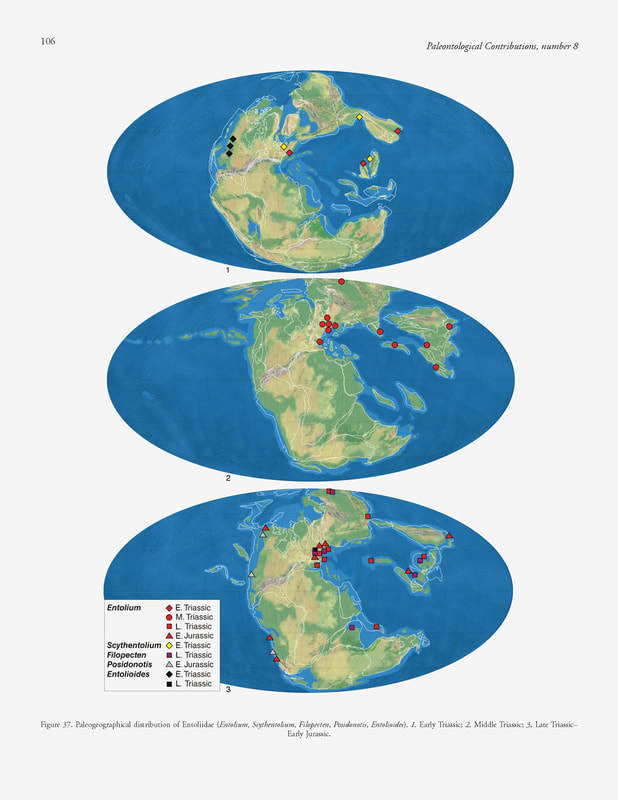
S. Ros-Franch, A. Márquez-Aliaga & S. E. Damborenea, 2014, Comprehensive database on Induan (Lower Triassic) to Sinemurian (Lower Jurassic) marine bivalve genera and their paleobiogeographic record, figure 37.
|
Genus ENTOLIUM Meek, 1865, p. 478
Type species.— Pecten demissus Meek, 1865, p. 478.
Stratigraphic range.— Lower Triassic–Upper Cretaceous (Maastrichtian) (Allasinaz, 1972; Abdel-Gawad, 1986). Cox and others (1969) assigned it a Middle Triassic–Upper Cretaceous range. The youngest record is from the Maastrichtian (Abdel-Gawad, 1986) and the oldest one from the Lower Triassic (E. discites Schlotheim, 1820).
Paleogeographic distribution.— Cosmopolitan (Fig. 37). In the Boreal domain, in addition to the Triassic, it was also recorded in the Early Jurassic (Kurushin, 1990; Polubotko & Repin, 1990), but the specimens were not figured or discussed. Milova (1976) reported it from the Pliensbachian and Milova (1988) from the Toarcian of northeastern Russia.
Tethys domain: Early Triassic: Italy (Allasinaz, 1972; Neri & Posenato, 1985), China (C. Chen, 1982; S. Yang, Wang, & Hao, 1986); Middle Triassic: Hungary (Allasinaz, 1972; Szente, 1997), Germany (Bachmann, 1973; Hagdorn, 1995), Poland (Senkowiczowa, 1985); Anisian of Italy (Allasinaz, 1972; Posenato, 2008b), China (Gu & others, 1976; Sha, Chen, & Qi, 1990; J. Chen, 2003), Bulgaria (Tronkov & Damyanov, 1993), Bosnia and Yugoslavia (Allasinaz, 1972), northern Vietnam (Komatsu, Huyen, & Huu, 2010); Ladinian of Spain (Márquez-Aliaga, 1983, 1985; Márquez-Aliaga & Montoya, 1991; Budurov & others, 1991; López-Gómez & others, 1994), China (Gu & others, 1976), Malaysia (Tamura, 1973), Italy (Rossi Ronchetti, 1959; Allasinaz, 1972), northern Vietnam (Komatsu, Huyen, & Huu, 2010), Afghanistan (Farsan, 1972); Late Triassic: Carnian of the Alps (Allasinaz, 1966, 1972; Fürsich & Wendt, 1977; Hautmann, 2001b), Spain (Martín-Algarra, Solé de Porta, & Márquez-Aliaga, 1995), China (Gu & others, 1976; Sha, Chen, & Qi, 1990); Norian of China (Lu, 1981); Norian–Rhaetian of Iran (Hautmann, 2001b); Rhaetian of ?Tibet (“Lhasa Block”) (J. Yin & Grant-Mackie, 2005), Alps (Vöros, 1981; Hautmann, 2001b), Hungary (Vörös, 1981; Hautmann, 2001b), Italy (Sirna, 1968); Early Jurassic: Hettangian of the Alps (Johnson, 1984), Germany and France (Vörös, 1971; Johnson, 1984), Vietnam (Vu Khuc & Huyen in Sato & Westermann, 1991). Circumpacific domain: Early Triassic: Olenekian of Japan (Nakazawa, 1961; Hayami, 1975; Kashiyama & Oji, 2004); Middle Triassic: Japan (Hayami, 1975; Tamura & others, 1978); Early Jurassic: Hettangian of Chile (Hillebrandt, 1990); Hettangian–Sinemurian of Canada (Aberhan, 1998a; Aberhan, Hrudka, & Poulton, 1998), Chile (Aberhan, 1993, 1994a); Sinemurian of Japan (Hayami, 1975), Canada (Poulton, 1991). Austral domain: Early Jurassic: Sinemurian of Argentina (Damborenea, 2002a; Damborenea & Manceñido, 2005b). Boreal domain: Middle Triassic: Anisian of Siberia (Dagys & Kurushin, 1985); Late Triassic: Carnian of Primorie (Kiparisova, 1972); Norian of Siberia (Okuneva, 1985). Paleoautoecology.— B, E, S, Un, FaM-Sed; R-Sw. Young specimens of Entolium s.s. had a byssal notch, but this was lost in adult stages. The life habit also probably changed from epibyssate in early stages to reclined in the upper ones. The low convexity of the shell, its reduced thickness, its circular outline, and the wide umbonal angle are characteristics that indicate it could have been a good swimmer, like some living pectinids (Johnson, 1984). However, in our provisional, broad concept of Entolium, there are some species that retained the byssal notch until the adult stages and therefore were epibyssate their entire lives.
Mineralogy.— Bimineralic (Carter, 1990a, p. 257). The family Entoliidae is characterized by bimineralic mineralogy (Carter, 1990a), with different microstructure types in juveniles and adults (homogeneous, simple prismatic, foliated, and fibrous prismatic), and a cross-lamellar inner shell layer (see Allasinaz, 1972; Waller, 1978; Johnson, 1984, among others, for more information).
ROS-FRANCH, S., A. MÁRQUEZ-ALIAGA & S. E. DAMBORENEA. 2014. Comprehensive database on Induan (Lower Triassic) to Sinemurian (Lower Jurassic) marine bivalve genera and their paleobiogeographic record. Paleontological Contributions, 8: 3-219, figs. 1-61. [p. 107]
|

Entolium kolymaense Kiparisova; L. G. Milova, 1976, Stratigraphy and bivalve molluscs of Triassic
and Jurassic deposits of northern Priokhoyta, plate 6, figures 2, 3; Entolium kolymaense pachyfibrosum Milova subsp. nov., figures 4, 5. |
«Because of the new distinction between the family Entolioididae and the entoliid subfamily Entoliinae, the age of the oldest member of the Entoliinae is uncertain pending the examination of shell microstructure. Tentatively, the oldest known species appears to be Entolium kolymaense Kiparisova, 1947 from the Upper Triassic (Carnian to Norian–Rhaetian) of south-eastern Siberia, based on specimens illustrated by Milova (1976: plate 6, figs 2–5). The youngest member of the Entoliinae appears to be Entolium orbiculare (Sowerby, 1817), which ranges into the early Late Cretaceous (Cenomanian) (Dhondt, 1971: 26; Kelly, 1984: 81).»
WALLER, T. R. 2006. Phylogeny of families in the Pectinoidea (Mollusca: Bivalvia): importance of the fossil record. Zoological Journal of the Linnean Society, 148: 313-342, figs. 1-12. [p. 328]
|
|
«Family ENTOLIIDAE Korobkov, 1960
Genus ENTOLIUM Meek, 1865 Type species. Original designation; Meek 1865, p . 478; Pecten demissus Phillips, "as illustrated by Quenstedt, 1858, p . 353, pl. 48, figs. 6, 7", i.e. Entolium demissum Meek, 1865 under Art. 70b of the Code [non Phillips; = Pecten disciforme Schübler in Zieten 1833, p. 69, pl. 53, fig. 2]; from the Aalenian of Germany.
Diagnosis. Byssal notch usually absent in all stages of development; margins closed laterally; incised ligamental area usually present and extending parallel to the hinge line of either side of the resilifer; cardinal crura variably developed, auricular crura present; inner layer of foliated calcite, outer layer radially fibrous.»
DUFF, K. L. 1978. Bivalvia from the English Lower Oxford Clay (Middle Jurassic). Palaeontographical Society. Monograph 553: 1-137, pls. 1-13. [p. 61]
|