Agerchlamys Damborenea, 1993
DAMBORENEA, S. E. 1993. Early Jurassic South American pectinaceans and circum-Pacific palaeobiogeography. Palaeogeography, Palaeoclimatology, Palaeoecology, 100: 109-123, figs. 1-4. [p. 115, 119]
|
«Agerchlamys n. gen.
This group (see Appendix and Fig. 4) includes finely ornamented species known from the circum-Pacific Late Triassic-Early Jurassic. The palaeogeographical distribution of this genus shows a distinctively bipolar pattern during the Pliensbachian (Fig. 2A), with records from northeast Russia (Milova, 1976), New Zealand (Marwick, 1953) and west-central Argentina (Damborenea, 1991, in press b). During the Late Triassic the distribution was similar and it has been found in Boreal (Kiparisova et al., 1966) and Austral regions (material seen at Otago University, New Zealand). Agerchlamys wunschae (Marwick) is found in New Zealand and the Andes only in very fine grained sediments, from marls to tuffaceous siltstones, but never in laminated dark mudstones. Although locally abundant in certain beds, it is otherwise not widespread and seems to have been a stenotopic species limited to very low-energy but well-oxygenated environments. It is generally associated with a variety of limid species and commonly occurs with sponge spicules. It is never associated with corals or thickshelled epifaunal bivalves. Agerchlamys wunschae was a byssally attached pectinid, as shown by the deep byssal notch and strong ctenolium (Fig. 4). Sponges could have provided a suitable substrate for this species.
[...]
Appendix: Description of Agerchlamys new genus
Superfamily: PECTINACEA Rafinesque 1815
Family: PECTINlDAE Rafinesque 1815
Subfamily: CAMPTONECTINAE Habe 1977
Genus: Agerchlamys n. gen.
Type species: Chlamys (Camtpochlamys) wunchae Marwick, 1953, from the Early Jurassic (Aratauran) of New Zealand and Pliensbachian of the Andes (type specimen figured in Marwick, 1953, plate 10, figs. 23, 24; Speden and Keyes, 1981, plate 13, figs. 2, 7; Fig. 4). A detailed description of the species will be provided elsewhere (see Damborenea, 1991).
Etymology: This genus is dedicated to Professor Derek Ager. It is especially appropriate taking into account Derek's interest in palaeobiogeography of benthonic invertebrates during the Mesozoic (Ager, 1986).
Diagnosis
Medium to large-sized pectinids, with sub-equivalve and subequilateral thin, discoidal, compressed shells. Umbonal angle wider than 90º. Anterior auricle at least twice as long as the posterior. Right anterior auricle with deep ctenoliate byssal notch. Fine cancellate ornamentation comprises very numerous (more than 40) narrow, nearly straight radial costae, dense continuous comarginal lamellae and fine antimarginal striae over the whole shell surface. Disc and auricles equally ornamented. Inner shell surface almost smooth. Central triangular resilifer. one cardinal crus (or dorsal tooth) parallel to the hinge axis and one tooth-like crus (or resilial tooth) to each side of resilifer. No auricular crura. Included species
Chlamys (Camptochlamys) inspecta Kiparisova, 1966 (in Kiparisova et al., 1966, pp. 124- 126, plate 17, figs. 7-10, plate 18, fig. 1, 2) from the Carnian-Norian of Siberia. Chlamys (Camptochlamys) wunschae Marwick, 1953 (pp. 98, 99, plate 10, figs. 23, 24; Fig. 4) from the Pliensbachian of New Zealand and Argentina.
Chlamys (Camptochlamys) proprius Milova, 1976 (pp. 60, 61, plate 7, figs. 10, 11) from the Pliensbachian of Siberia.
To the above-mentioned species, perhaps Chlamys ex. gr. textoria (Schlot.) in Polubotko, 1968b (pp. 78, 79, plate 23, figs. 5, 6) from the Pliensbachian of Siberia could be added. Some northern European specimens, generally referred to Chlamys torulosi (Quenstedt) are superficially similar (see for instance Staesche, 1926, p. 33, plate 1, fig. 1), though nobody reported antimarginal ornamentation on these forms. In his revision of Jurassic European pectinids, Johnson (1984, p. 170) includes them all within Chlamys textoria, accepting a very wide range in the number of radial costae. It is interesting to point out that C. wunschae was also doubtfully included in Johnson's (1984) synonymy of Chlamys lextoria, but he later states (p. 173) that the antimarginal ornamentation "probably serves to distinguish the species."
Also some Middle Jurassic material from European regions may also be related to this taxon, but a full analysis must wait until more information about them becomes available.
Discussion
Species of this taxon have been referred either to Chlamys or to Camptochlamys in the past, though it has been regarded as a distract group. According to the general shell shape, hinge details, presence of antimarginal ornamentation and type of ornamentation pattern the group of A. wunschae can be referred to the Camptonectinae in the sense of Waller and Marincovich (1992) and has affinities to Camptochlamys. The group has, nevertheless, a more delicate and wavy nature of the concentric lamellae that, together with the higher density of both radial and comarginal elements of the ornamentation, gives the shell a finer appearance if compared with the type species of Camptochlamys. The numerous, regularly spaced, fine comarginal lamellae, are norreally stronger and convex outwards on the interspaces than on the costae in Agerchlamys. The shell is also thinner in the group of species under consideration, which also have radial ornamentation on the auricles and a gently scalloped inner shell margin.
This group of species could also be compared to some finelyornamented pectinids referred to Chlamys or Lyriochlamys by most authors, but in this case, the Camptonectes-like shape of the shell, the wide umhonal angle, the presence of costae instead of plicae, of antimarginal striae over the whole shell and of continuous comarginal lamellae are the characters which prevent assignlnent to this genus. Relations with a Boreal stock of finely ribbed pectinids, which flourished from Late Triassic to mid-Jurassic times, is still unclear. These are traditionally referred to Chlamys. The most conspicuous member of this group is C. mojsisovicsi Kobayashi and lchikawa (1949: see also Hayami, 1975) fiom the Late Triassic of Japan, Siberia and western North America. Though the ornamentation pattern is comparable, species of this group seem to lack antimarginal ornamentation.
Distribution
Agerchlamys ranges in age from the Late Triassic to the Toarcian of the Boreal and Austral Pacific regions (Fig. 2A).» SUSANA ESTER DAMBORENEA, 1993
|
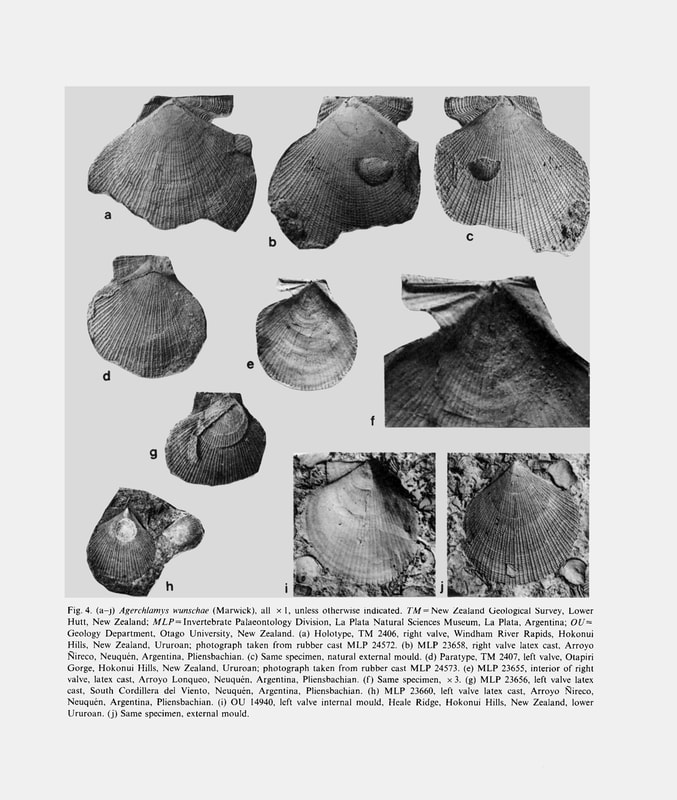
Agerchlamys wunschae (Marwick); S. E. Damborenea, 1993, Early Jurassic South American pectinaceans and circum-Pacific palaeobiogeography, figures 4a-4j.
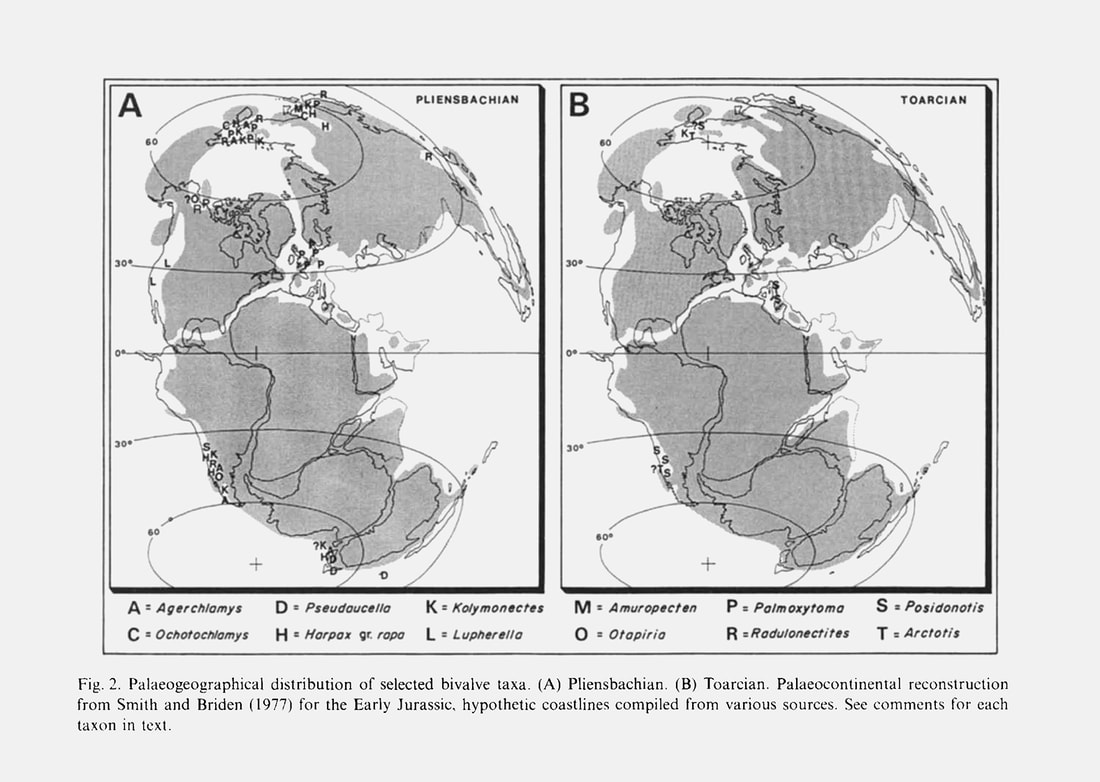
S. E. Damborenea, 1993, Early Jurassic South American pectinaceans and circum-Pacific palaeobiogeography, figure 2.
|
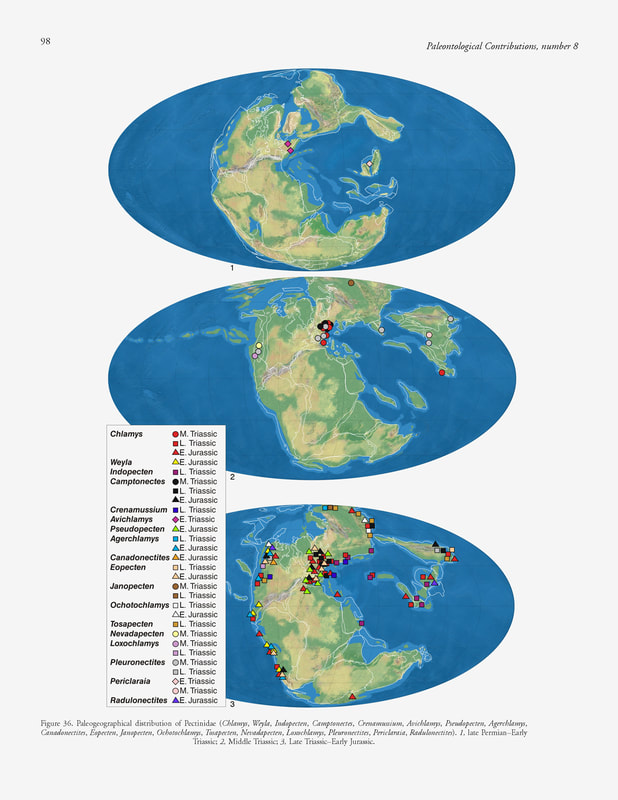
S. Ros-Franch, A. Márquez-Aliaga & S. E. Damborenea, 2014, Comprehensive database on Induan (Lower Triassic) to Sinemurian (Lower Jurassic) marine bivalve genera and their paleobiogeographic record, figure 36.
|
Genus AGERCHLAMYS Damborenea, 1993, p. 119
Type species.— Chlamys (Camptochlamys) wunschae Marwick, 1953, p. 98.
Stratigraphic range.— Upper Triassic (Carnian)–Lower Jurassic (Toarcian) (Damborenea, 1993, 2002a). Damborenea (1993) proposed Agerchlamys, including several previously described species referred to other genera (see Damborenea, 1993, p. 120, and Damborenea, 2002a, p. 66, for species listed). These species were recorded from the Carnian to the Toarcian, and the author indicated the possibility that the genus may be present up to the Middle Jurassic.
Paleogeographic distribution.— Circumpacific, Austral, and Boreal (Fig. 36). Agerchlamys was distributed through the Austral and Boreal domains and also in the Circumpacific, but always at high latitudes. In the Austral domain (Argentina and New Zealand), it was reported primarily from Pliensbachian beds (Marwick, 1953; Damborenea, 1993, 2002a).
Circumpacific domain: Early Jurassic: Hettangian of Chile (Aberhan, 1994a), Oregon (United States) (D. G. Taylor & Guex, 2002), British Columbia (western Canada) (Wignall & others, 2007); Hettangian–Sinemurian of Canada (Aberhan, 1998a, 1998b), Sonora (Mexico) (Scholz, Aberhan, & González-León, 2008). Austral domain: Early Jurassic: Hettangian–Sinemurian of Argentina (Damborenea, 2002b; Damborenea & Manceñido, 2005b). Boreal domain: Late Triassic: Carnian–Norian of Siberia (Kiparisova, Bychkov, & Polubotko, 1966). Paleoautoecology.— B, E, S, Epi, Sed; By. Due to the presence of a deep byssal gape below the right anterior auricle and a strong ctenolium, it was epibyssate (Damborenea, 1993, 2002a). Although it has a wide umbonal angle, the auricles are of different sizes, so it is not believed that it could swim.
Agerchlamys is usually found in low-energy and well-oxygenated environments, and associated with sponges and other bivalves, especially limids (Damborenea, 2002a). Mineralogy.— Bimineralic (Carter, 1990a, p. 260). There are no data about Agerchlamys shell. We assume bimineralic mineralogy, as in the other members of the family Pectinidae (Carter, 1990a).
ROS-FRANCH, S., A. MÁRQUEZ-ALIAGA & S. E. DAMBORENEA. 2014. Comprehensive database on Induan (Lower Triassic) to Sinemurian (Lower Jurassic) marine bivalve genera and their paleobiogeographic record. Paleontological Contributions, 8: 3-219, figs. 1-61. [p. 103]
|