Eopecten Douvillé, 1897
DOUVILLÉ, H. 1897. Essai de classification systématique des pectinidés. Bulletin de la Société Géologique de France [3e Série], 25: 202-205. [p. 203]
|
«3° A partir des temps secondaires, les Pectinidés constituent plusieurs groupes distincts qui évoluent d'une manière indépendante:
Premier groupe. — La surface est ornée de grosses côtes, souvent inégales et quelquefois costulées. Les espèces les plus anciennes, souvent confondues à tort avec les Hinnites, affectent la forme Semipecten; on peut les désigner sous le nom de Eopecten n. gen. (type H. tuberculatus Goldf.), Trias, Jura, et Crét. inf.; ce type donne naissance d'un côté à une forme Hinnites (G. Terquemia) et de l'autre à des formes demi-libres, G. Pecten (Pollux, valoniensis) et libres, G. Pseudopecten (aequivalvis), de la forme Chlamys. HENRI DOUVILLÉ, 1897
|

Spondylus tuberculosus nobis; A. Goldfuss, 1833-1840, Petrefacta Germaniae, plate 105, figures 2a, 2b.
|
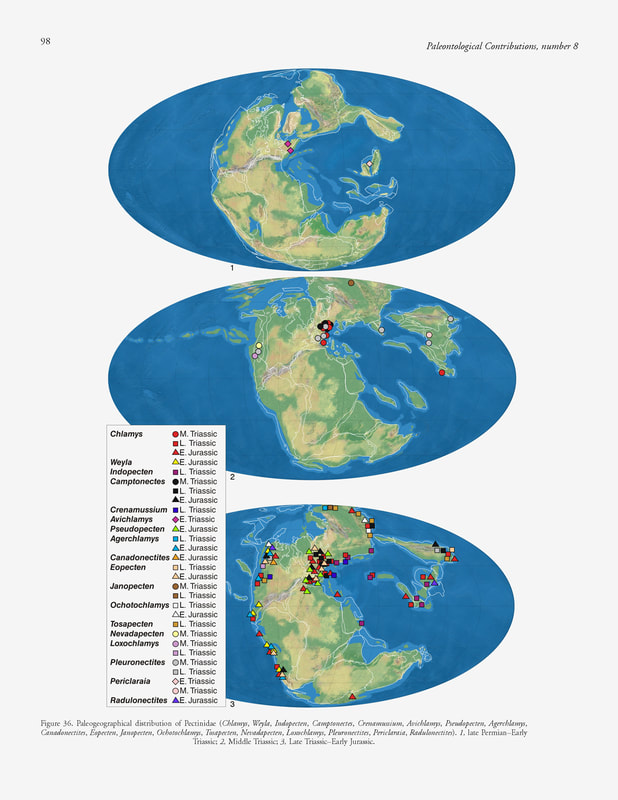
S. Ros-Franch, A. Márquez-Aliaga & S. E. Damborenea, 2014, Comprehensive database on Induan (Lower Triassic) to Sinemurian (Lower Jurassic) marine bivalve genera and their paleobiogeographic record, figure 36.
|
«Genus EOPECTEN Douvillé, 1897, p. 203
Type species.— Hinnites tuberculatus Goldfuss (errore pro Spondylus tuberculosus Goldfuss), 1835 in 1833–1841, p. 93.
Remarks.— See Johnson (1984, p. 149) and Damborenea (1987b, p. 198) for a discussion about synonymy and the problems related to this genus since its proposal.
Stratigraphic range.— Upper Triassic (Carnian)–Lower Cretaceous (Albian) (Hayami, 1975; Johnson, 1984). Cox and others (1969) assigned it a Jurassic–Lower Cretaceous (Albian) range. Although it seems fairly accepted that it appeared in the Early Jurassic, according to the literature, there are several records from the Carnian of Japan (Kobayashi & Ichikawa, 1949b; Nakazawa, 1952; Hayami, 1975) and from the Norian of Chile (Hayami, Maeda, & Ruiz-Fuller, 1977) that no other author except Hallam (1981) considered. Middle Triassic specimens assigned by Allasinaz (1972) to Radulonectites should be allocated to Eopecten instead (Damborenea, 2002a, p. 61). If Radulonectites? flagellum (Stoppani, 1858 in 1858–1860), described by Allasinaz (1972, p. 331), is assigned to Eopecten, the genus was present from Ladinian times. Another species that was referred to Eopecten, originally proposed as Monotis albertii Goldfuss, 1835 in 1833–1841 (Diener, 1923), was reported from the Lower and Middle Triassic of Europe, but it is currently included in Leptochondria (Waller & Stanley, 2005, p. 34).
Paleogeographic distribution.— Tethys, Circumpacific, and Austral (Fig. 36). According to the published records, it seems that Eopecten originated in the Late Triassic of Japan, and then it migrated to the western Tethys (Europe) and eastern Paleopacific.
Tethys domain: Early Jurassic: Hettangian of England (Liu, 1995), Belgium and Germany (Johnson, 1984); Sinemurian of Portugal and Spain (Liu, 1995), Germany (Johnson, 1984). Circumpacific domain: Late Triassic: Carnian of Japan (Kobayashi & Ichikawa, 1949b; Nakazawa, 1952; Hayami, 1975); Early Jurassic: Hettangian of Chile (Aberhan, 1994a); Hettangian–Sinemurian of Canada (Aberhan, 1998a); Sinemurian of Canada (Poulton, 1991). Austral domain: Early Jurassic: Hettangian–Sinemurian of Argentina (Riccardi & others, 1991; Damborenea, 2002a; Damborenea & Manceñido, 2005b). Paleoautoecology.— B, E, S, Epi-C, Sed; By-C. Johnson (1984) and Harper and Palmer (1993) analyzed the mode of life of different species of Eopecten. The last authors concluded that some species could live cemented to the substrate, while others were epibyssate during most of their life.
Mineralogy.— Bimineralic (Carter, 1990b, p. 388; Harper & Palmer, 1993, p. 67). The shell of Eopecten had a foliated outer shell layer, both in the left valve (Carter, 1990a) and in the right one (Harper & Palmer, 1993), and aragonitic middle and inner shell layers with cross-lamellar microstructure (Carter, 1990a).»
ROS-FRANCH, S., A. MÁRQUEZ-ALIAGA & S. E. DAMBORENEA. 2014. Comprehensive database on Induan (Lower Triassic) to Sinemurian (Lower Jurassic) marine bivalve genera and their paleobiogeographic record. Paleontological Contributions, 8: 3-219, figs. 1-61. [p. 102, 103]
|

Eopecten spondyloides (Roemer, 1836); A. L. A. Johnson, 1984, The palaeobiology of the bivalve families Pectinidae and Propeamussiidae in the Jurassic of Europe, plate 5, figures 9-14.
|
«Genus EOPECTEN DOUVILLÉ 1897
(Synonyms etc. Velata Quenstedt 1856 [non Griffith and Pidgeon 1934, obj.]
Velopecten Philippi 1899 [pro Velata Quenstedt 1856, non Velates Montfort 1810] Velatopecten Rollier 1906 [nom. van.]) Type species. OD; DOUVILLÉ 1897, p. 203; Hinnites tuberculatus Goldfuss errore pro Spondylus tuberculosus Goldfuss 1836, p. 93, pl. 105, fig. 2; Aalenian/Bajocian, Swabia.
AMENDED DIAGNOSIS
Medium sized to large, acline, more or less irregulär in outline at all stages of growth, some specimens irregularly puckered or with allomorphic ornament; inequivalve, with LV convex and RV flat or concave; left anterior wing large, indistinctly demarcated, right anterior auricle elongate, deep subauricular notch with ctenolium below it; posterior wing rather small; RV with narrow, obtusely triangulär cardinal area with deep, narrowly triangulär pit below beak corresponding to resilium (same in LV); interior of RV in some specimens with blunt oblique internal ridge originating near resilifer; ornament of Striae and costae of varying strengths. L. Jur. (Hettang.)-L. Cret. (Alb.), cosmop.»
JOHNSON, A. L. A. 1984. The palaeobiology of the bivalve families Pectinidae and Propeamussiidae in the Jurassic of Europe. Zitteliana, 11: 1-235, pls. 1-11. [p. 149]
|