Catillopecten natalyae Kamenev, 2017
KAMENEV, G. M. 2018. Four new species of the family Propeamussiidae (Mollusca: Bivalvia) from the abyssal zone of the northwestern Pacific, with notes on Catillopecten squamiformis (Bernard, 1978). Marine Biodiversity, 48: 647-676, figs. 1-16 (published online: 21 November 2017). [p. 664, figs. 12-14]
2017 Catillopecten natalyae Kamenev, 2017
G. M. Kamenev, 2018, figures 12-14.
|
«Cyclopecten sp. 2: Kamenev 2015, p. 191.
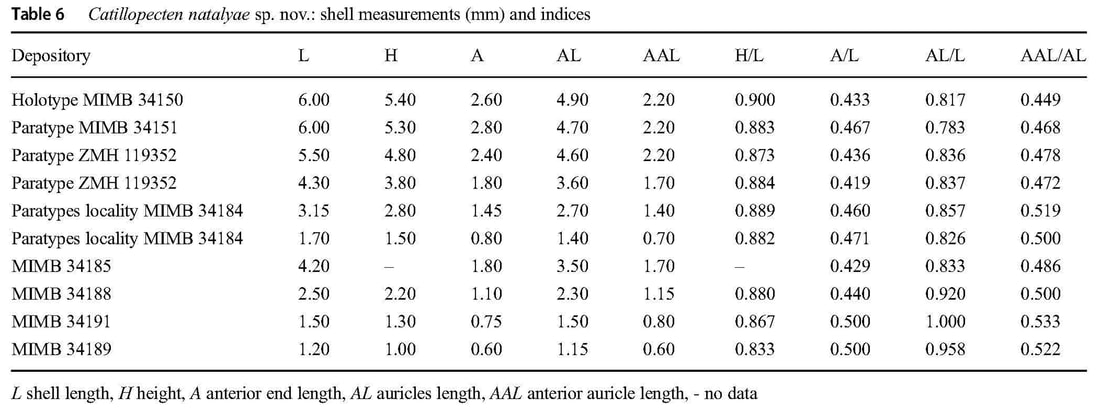
Type material and locality: Holotype (MIMB 34150), abyssal plain adjacent to Kuril-Kamchatka Trench, Pacific Ocean (43°02,31′ N, 152°59,16′ E), 5,222 m, box corer, Coll. G. M. Kamenev, 16-VIII-2012 (RV Sonne, cruise no. 223, stn. 7-4); paratypes (3) (paratype, MIMB 34151; paratypes (2), ZMH 119352), abyssal plain adjacent to Kuril-Kamchatka Trench, Pacific Ocean (43°58,35′N, 157°19,74′E – 43°58,62′N, 157°18,15′E), 5,407-5,418 m, epibenthic sledge, Coll. A. Brandt, 30-VII-2012 (RV Sonne, cruise no. 223, stn. 1-11).
Other material examined: Four damaged specimens (MIMB 34184) from paratypes locality; 1 specimen (MIMB 34185), abyssal plain adjacent to Kuril-Kamchatka Trench, Pacific Ocean (43°35,50′N,153°57,89′E – 43°34,30′N, 153°58,18′E), 5,378-5,376 m, epibenthic sledge, 11-VIII-2012 (RV Sonne, cruise no. 223, stn. 5-9); 1 heavily damaged young specimen (MIMB 34186), abyssal plain adjacent to Kuril-Kamchatka Trench, Pacific Ocean (43°02,78′N, 152°59,30′E – 43°01,65′N, 152°58,45′E), 5,217-5,223 m, epibenthic sledge, 17–18-VIII-2012 (RV Sonne, cruise no. 223, stn. 7-10); 1 heavily damaged specimen (MIMB 34187), abyssal plain adjacent to Kuril-Kamchatka Trench, Pacific Ocean (42°14,69′N, 151°44,05′E – 42°14,26′N, 151°42,49′E), 5,127 m, epibenthic sledge, 20-VIII-2012 (RV Sonne, cruise no. 223, stn. 8-9); 1 shell of young specimen (MIMB 34188), abyssal plain adjacent to Kuril-Kamchatka Trench, Pacific Ocean (42°14,73′N, 151°44,38′E – 42°14,34′N, 151°42,95′E), 5,112-5,126 m, epibenthic sledge, 21-VIII-2012 (RV Sonne, cruise no. 223, stn. 8-12); 2 specimens (MIMB 34189), abyssal plain adjacent to Kuril-Kamchatka Trench, Pacific Ocean (40°35,49′N, 150°59,92′E – 40°34,25′N, 150°59,91′E), 5,399-5,398 m, epibenthic sledge, 23–24-VIII-2012 (RV Sonne, cruise no. 223, stn. 9-9); 1 heavily damaged young specimen (MIMB 34190), abyssal plain adjacent to Kuril-Kamchatka Trench, Pacific Ocean (41°11,70′ N, 150°05,56′E – 41°12,99′N, 150°05,59′E), 5,250-5,245 m, epibenthic sledge, 27–28-VIII-2012 (RV Sonne, cruise no. 223, stn. 10-12); 1 young specimen (MIMB 34191), abyssal plain adjacent to Kuril-Kamchatka Trench, Pacific Ocean (40°13,10′N, 148°06,45′E – 40°12,10′N, 148°05,53′E), 5,351-5,348 m, epibenthic sledge, 31-VIII-2012 (RV Sonne, cruise no. 223, stn. 11-12). Total of 11 specimens and 1 shell of young specimen.
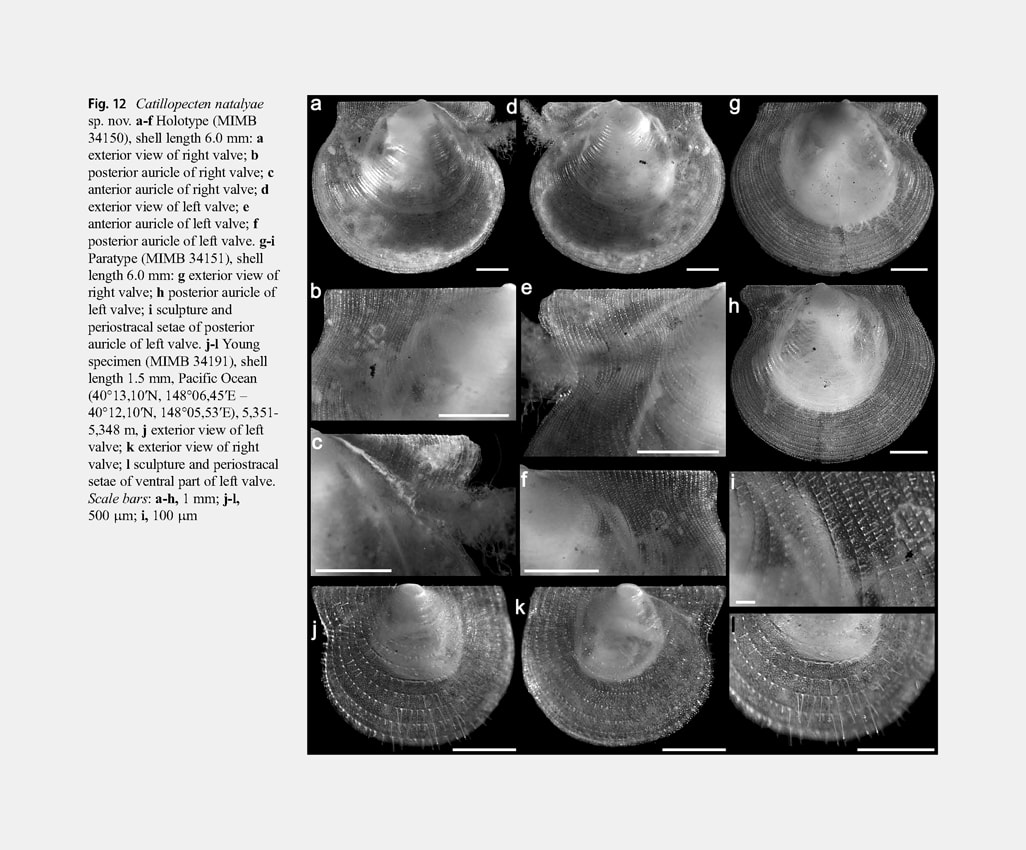
Description: Shell small (to 7.1 mm in length), very thin and fragile, translucent, weakly inflated, D-shaped, slightly longer than higher (H/L = 0.833-0.900), inequilateral (beaks slightly anterior to midline, A/L = 0.419-0.500), inequivalve (in large specimens (shell length more than 5.0 mm) right valve convex and left valve only slightly more convex than right). Dorsal shell margin straight. Anterior, posterior and ventral shell margins smoothly rounded. Auricles very large (AL/L = 0.783-1.000), unequal in length (anterior auricles in adults shorter than posterior, AAL/AL = 0.449-0.478) and dissimilar in shape; anterior auricles slightly twisted to the left. Umbonal angle 120°.
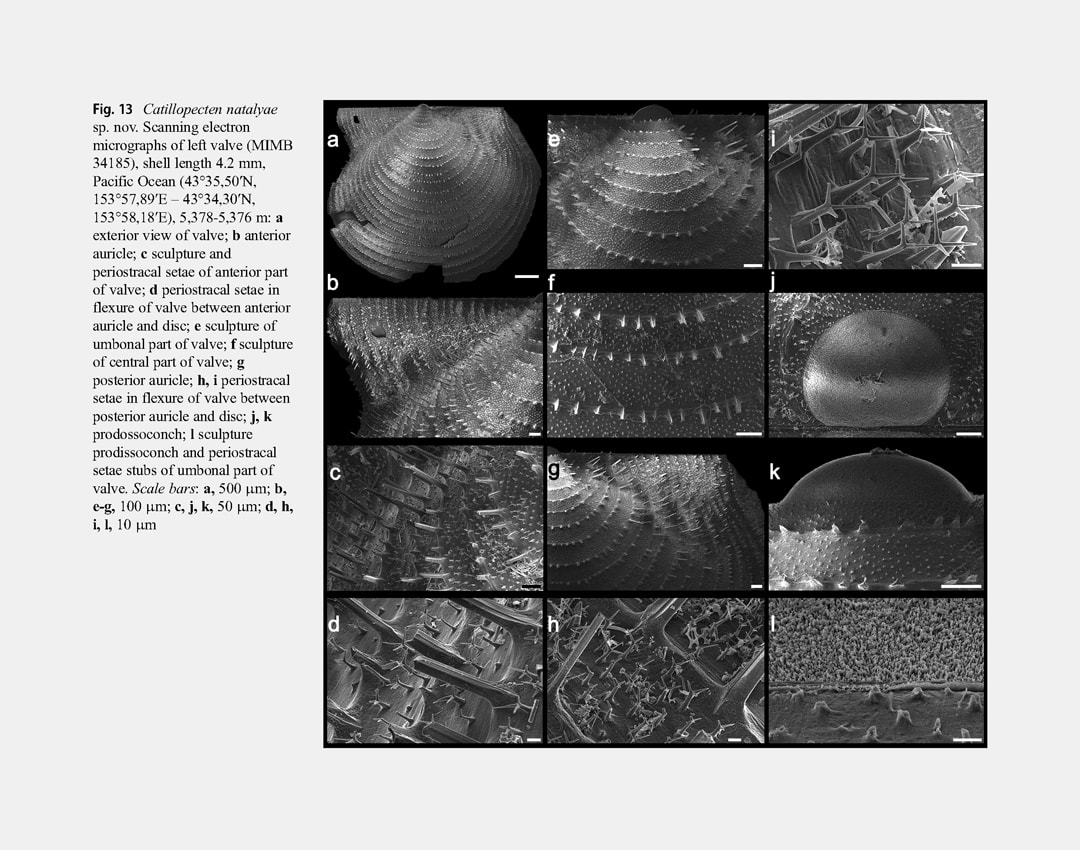
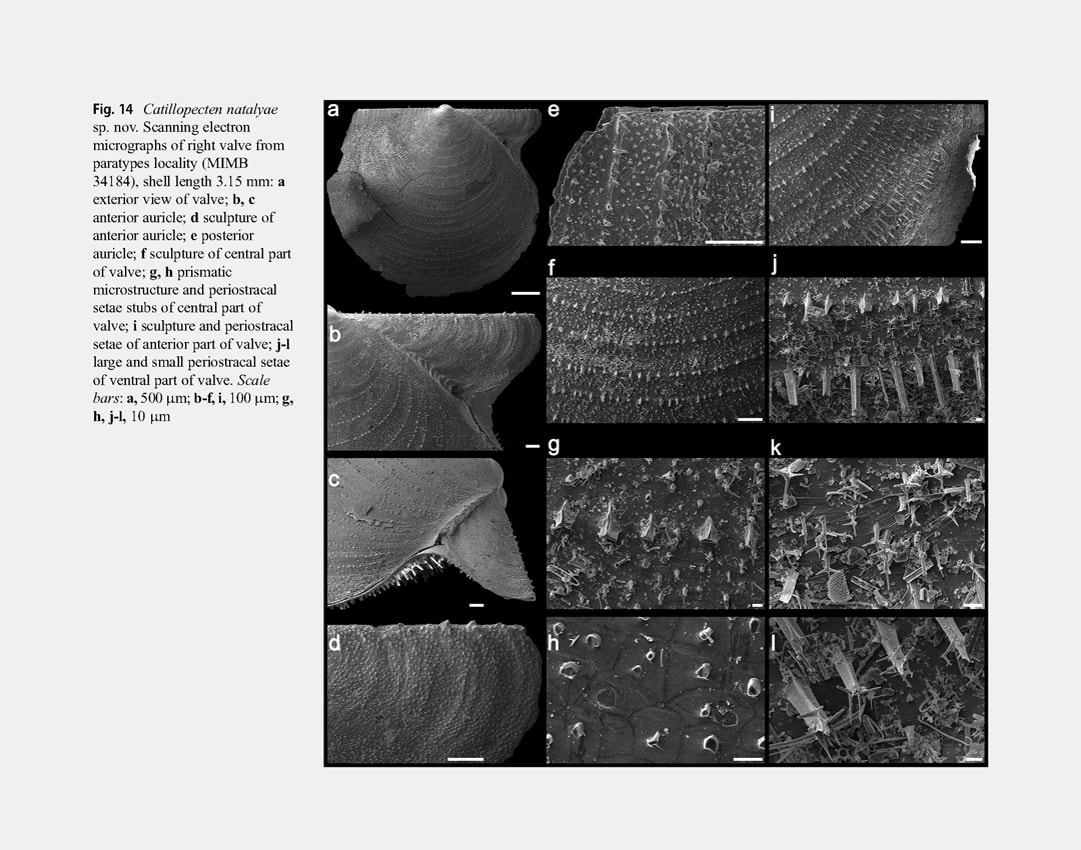
Right valve disc with weak, indistinct, regular, broad, commarginal undulations, sculptured with closely spaced, regular, very thin and fine, commarginal riblets (interval of 100-120 μm between riblets in central part of disc). Fine commarginal riblets with closely spaced, three-edged periostracal setae broken off at different height (the highest stubs and almost undamaged setae at ventral valve margin); only bases and very short stubs of broken periostracal setae near beak and on auricles (Fig. 14f, i). Each edge of longest broken three-sided setae dichotomously branching at ventral
valve margin into 2 long, rounded, pointed processes diverging at an angle of 100-110° (Fig. 14l). In intervals between commarginal riblets with large, three-edged, broken setae are numerous undamaged and broken short (to 10 μm in height), triangular-rounded, periostracal setae with apices branching into 3 long, rounded, pointed processes diverging at 120° (Fig. 14j, k). Outer prismatic layer consisting of commarginal arrays of high prisms (70-80 μm in height); commarginal riblet with periostracal setae corresponding to each row of high prisms; between arrays of high prisms are bands of 4-6 sided small prisms (7-12 μm in height) 6-7 prisms wide (Fig. 14g, h). Anterior auricle with rounded anterior end, sharply demarcated from shell disc by a byssal fasciole. Surface of auricle with very weak, irregular, commarginal undulations and commarginal rows of short stubs of broken long periostracal setae with numerous stubs of short periostracal setae between these commarginal rows. Byssal notch moderately deep, sharp. Byssal fasciole narrow, folded. Posterior auricle continuous with shell disc, similarly sculptured as a valve disc. Left valve disc not commarginally undulated and similarly sculptured as a right valve disc. Stubs broken long periostracal setae on commarginal riblets of shell disc higher than on right valve, reaching maximum length at valve margins, in flexure of valve between anterior auricle and disc, and on posterior auricle (Fig. 13b–g-i). Anterior auricle triangular, poorly demarcated from shell disc by a distinct, wide, radial flexure; byssal sinus shallow. Posterior auricle continuous with shell disc. Anterior and posterior auricles similarly sculptured as a shell disc. Prodissoconch large (length 270-290 μm), D-shaped, convex, distinct, sharply separated from shell disc, with brushlike surface of very closely spaced, short columns. Variability: In smaller specimens (shell length less than 3 mm) compared to larger ones, the right valve is flattened, the shell is higher (H/L = 0.883-0.900), the total length of auricles is relatively greater (equal or almost equal to the shell length, AL/L = 0.824-1.000), the anterior auricle length is equal or greater than the posterior auricle length (AAL/AL = 0.500-0.583), the anterior auricle projects beyond the anterior shell margin, the long periostracal setae on commarginal riblets of valves are less damaged and their stubs are longer (especially on the left valve) (Fig. 12j-l).With increasing shell size, the length of stubs of periostracal setae decreases and in the largest specimens they become very short and weakly noticeable. In large specimens, the variability of shell shape and proportions is insignificant. In some specimens (e.g., in the holotype), the posterior auricle is slightly separated, compared to other specimens in which it is completely continuous
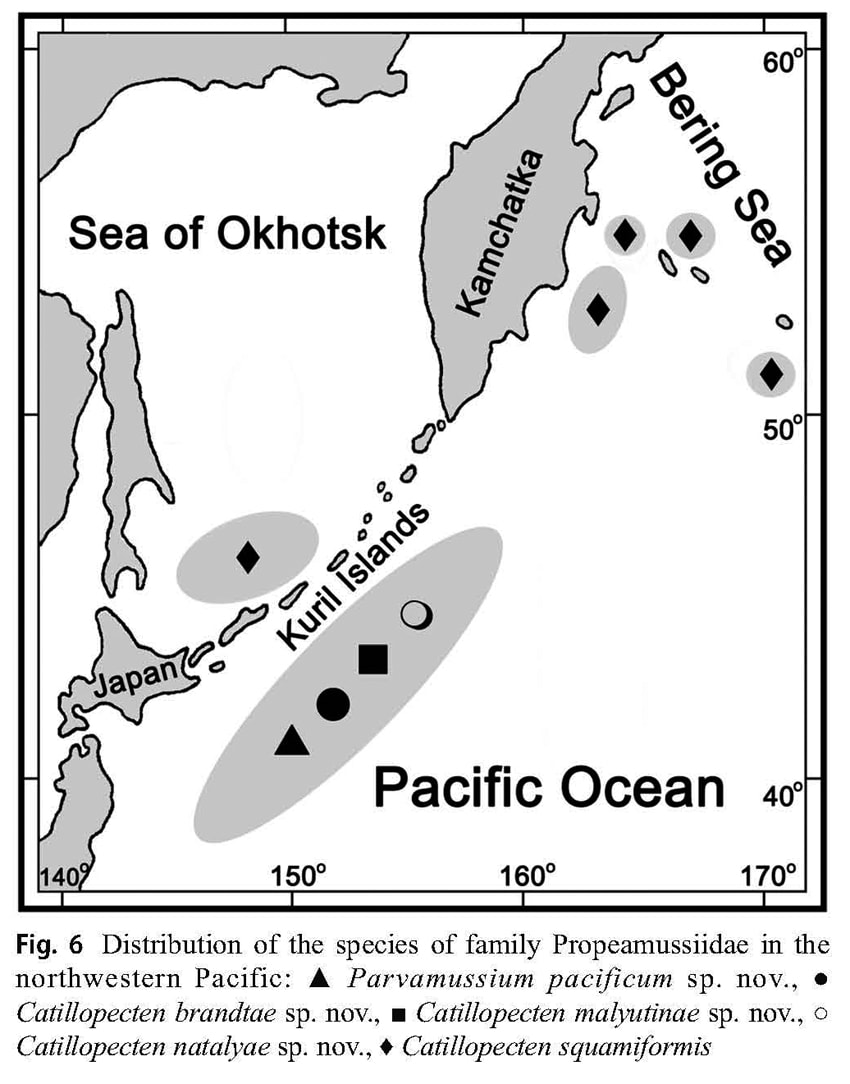
with the shell disc. Distribution and habitat (Fig. 6): This species was recorded at the abyssal plain adjacent to the Kuril-Kamchatka Trench (northwestern Pacific) at the latitudes from northern Honshu (Japan) (40°13,10′N, 148°06,45′E – 40°12,10′N, 148°05,53′E) to the southern Kuril Islands (43°58,35′N, 157°19,74′E – 43°58,62′N, 157°18,15′E) at 5,112-5,418 m depth (bottom temperature (6-8 m above bottom) 1.5-1.6 °C, salinity 34.7‰, oxygen 7.71-7.72 ml/l).
Comparisons: Catillopecten natalyae sp. nov. is easily distinguished from all species of the genus Catillopecten in having two types of periostracal setae on the shell: long, threeedged, located on commarginal riblets and short, with three apical processes, located in intervals between commarginal riblets (Table 3). In addition, only in this species commarginal undulations are present only on one valve and the prodissoconch has a brushlike surface of microscopic, closely set, short columns. Moreover, C. natalyae sp. nov. differs from most species of the genus Catillopecten in its D-shaped shell with very large auricles, the total length of which is almost equal to the shell length, and in having a triangular posterior auricle and byssal sinus on the left valve. Apart from C. natalyae sp. nov., only C. brandtae sp. nov. and C. micaceus have a D-shaped shell with very large auricles; however, they have no commarginal riblets and periostracal setae. Periostracal setae are only present in C. malyutinae sp. nov. In contrast to C. natalyae sp. nov., the periostracal setae in this species have a different structure and form antimarginal rows.
Derivatio nominnis: The species name honors my wife Natalya V. Kameneva, Head of the MIMB and a permanent and reliable assistant in my scientific work.
Remarks
Сatillopecten natalyae sp. nov.—as well as C. malyutinae sp. nov. and C. brandtae sp. nov.—is, to all appearances, widely distributed in the northwestern Pacific. It has been found from Honshu to the southern Kuril Island at most of the stations of the KuramBio expedition. All stations were conducted at depths greater than 5,000 m. Сatillopecten natalyae sp. nov. may occur in deeper waters not only at the abyssal plain but also in the hadal zone of the Kuril-Kamchatka Trench. Probably, it is more widely distributed and forms denser populations at the abyssal plain of the northwestern Pacific. It cannot be excluded that a part of specimens was destroyed when collecting and washing samples, because this species has a very thin and fragile shell.
A distinctive feature of С. natalyae sp. nov., as well as C. malyutinae sp. nov., is the presence on the shell of closely spaced periostracal setae, which are better preserved on the left upper valve. Similar broken periostracal setae of different size on commarginal riblets and in intervals between them are present on the shell of Cyclopecten ambiannulatus Schein, 1989 (Schein 1989). This is the deepest-water species of the genus Cyclopecten, which is widely distributed in the Atlantic at 800-4,900 m depth (Schein 1989; Olabarria 2005; Allen 2008; Dijkstra et al. 2009; Dijkstra and Janssen 2013). It is likely that the presence of periostracal setae on the shell is an adaptation in some species of the family Propeamussidae for life at maximum depths of abyssal plains. It is generally thought that most propeamussiids are free living, mobile carnivores (Knudsen 1979; Waller 1984; Allen 2008). However, in all probability, C. natalyae sp. nov. is a byssate species. One of the examined specimens of C. natalyae sp. nov. had retained a well-developed byssus with which it can attach itself to various bottom substrates. Waller (1984) supposed that most species of the living species of the family Propeamussiidae, including members of the genus Catillopecten, must be byssate, because they have a deep byssal notch throughout ontogeny.» GENNADY M. KAMENEV, 2017
|