Pecten novaezelandiae Reeve, 1852
REEVE, L. A. 1852-1853. Monograph of the genus Pecten. In: L. A. Reeve (Ed.), 1843-1878, Conchologia Iconica; or illustrations of the shells of molluscous animals, vol. 8. [unnumbered pages], pls. 1-35. London. [sp. 36, pl. 8, fig. 36]
1835 Pecten laticostata Gray, 1835
1852 Pecten novaezelandiae Reeve, 1852
1930 Notovola tainui Finlay, 1930
1930 Notovola marwicki Finlay, 1930
1951 Pecten toi Fleming, 1951
1951 Pecten kupei Fleming, 1951
1951 Pecten novazelandiae rakiura Fleming, 1951
1953 Pecten tainui aotea Fleming, 1953
1957 Pecten benedictus tepungai Fleming, 1957
1957 Pecten benedictus zeehaenae Fleming, 1957
1961 Pecten sulcicostatus var. casa van Bruggen, 1961
1852 Pecten novaezelandiae Reeve, 1852
1930 Notovola tainui Finlay, 1930
1930 Notovola marwicki Finlay, 1930
1951 Pecten toi Fleming, 1951
1951 Pecten kupei Fleming, 1951
1951 Pecten novazelandiae rakiura Fleming, 1951
1953 Pecten tainui aotea Fleming, 1953
1957 Pecten benedictus tepungai Fleming, 1957
1957 Pecten benedictus zeehaenae Fleming, 1957
1961 Pecten sulcicostatus var. casa van Bruggen, 1961

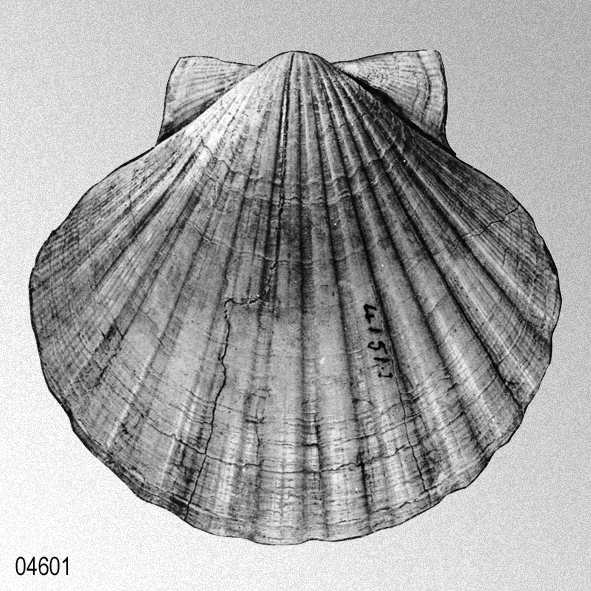
L. A. Reeve, 1852, plate 8.
|
«Pect. testa subelongato-orbiculari, aequilaterali, inaequivalvi; valvis concentrice minutissime striatis; sinistra plana, prope umbonem concaviuscula, in valva dextra subimmersa, costis subangustis convexis quatuordecim nitide radiata, albicante, spadiceo-rufo tincta; dextra expanso-convexa, costis quindecim sublatioribus radiata, alba; auriculis aequalibus, sinistrorsum subinclinatis.
THE NEW ZEALAND PECTEN. Shell somewhat elongately orbicular, equilateral, inequivalve; valves very minutely concentrically striated; left valve flat, rather concave near the umbo, a little immersed in the right valve, neatly rayed with fourteen rather narrow convex ribs, whitish, stained with fawn-red; right valve expandedly convex, rayed with fifteen rather broader ribs, white; ears equal, bent a little forwards towards the left valve. Hab. New Zealand; Hart. A beautifully symmetrical neatly-ribbed species, of a delicate subtransparent white, stained on the left valve with rich fawn-red.» LOVELL AUGUSTUS REEVE, 1852
|









































|
«Fleming (1951b, p. 129) selected the specimen in Lamarck’s collection (MHNG) as the lectotype of Pecten medius Lamarck, 1819 (junior secondary homonym of P. medius Bosc, 1802); this is a young specimen of Pecten maximus (Linné, 1758) (Fleming 1957, pl. 6, fig. 3, 4). Rosalie de Lamarck’s annotations on Lamarck’s copy of Animaux sans vertèbres (in MHNG) indicate that Lamarck saw only one specimen of Pecten medius, and Lamarck (1819) stated of P. medius: “Habite…Mus.no.”, that is, the specimen was already in MNHN at that time, and he made no indication of having a specimen in his collection (“Mon cabinet”) (Y. Finet MHNG pers. comm.). Dijkstra (1994, p. 471) therefore correctly considered that only the specimen in MNHN is a syntype. He considered Fleming’s designation to be invalid, and selected the Lamarck syntype in MNHN as the lectotype of Pecten medius Lamarck. In fact, as Lamarck had only one specimen, and did not refer to earlier illustrations, the MNHN specimen seems best interpreted as the holotype. This is a specimen of the P. meridionalis Tate, 1887 form of P. fumatus. Although P. medius was thought by several early authors to apply to P. novaezelandiae, it is actually a senior synonym of P. fumatus, as well as a junior secondary homonym.
[...]
REMARKS: The earliest occurrence of Pecten in New Zealand previously has been considered (following Fleming 1957) to be the rare specimens first discovered by Professor Paul Vella in Upper Westmere Shellbed (Abbott & Carter 1999), c. 1.3 m above Kaikokopu Shellbed in the coastal exposure at Castlecliff, Wanganui Basin. Kamp & Turner (1990) demonstrated that this position lies just above the Brunhes-Matuyama polarity transition, in rocks deposited early in OIS 19 (0.78 Ma); the transition is truncated by the unconformity at the base of Kaikokopu Shellbed. This position is very useful for correlation in Wanganui Basin, as it identifies the Brunhes-Matuyama magnetic transition wherever the earliest Pecten specimens can be found. However, in eastern New Zealand, the marwicki phenotype is known considerably earlier at two widely separated sites: Maraetotara Formation, Cape Kidnappers; and Leader River on “Mendip Hills” Station, inland from Parnassus, N Canterbury. At both these sites, Pecten occurs below Potaka Tephra (Black 1992; Shane 1994; Warren 1995, fig. 21; Proust & Chanier 2004, fig. 4; P. J. J. Kamp University of Waikato pers. comm.) and so Pecten here is more than 1.0 m yr old (Shane 1994; Shane et al. 1996; Alloway et al. 2005; Pillans et al. 2005), that is, deposited in about OIS 29–31, or possibly significantly earlier. The locality with abundant P. novaezelandiae at Herepuru Stream, coast W of Matata, Bay of Plenty, also possibly is older than the Brunhes-Matuyama transition, as it underlies a tuff dated at 0.62 ± 0.03 Ma (Nairn & Beanland 1989, table 1) by an interval of unknown duration, but this locality is assigned here to “about OIS 15”.
The forms of Pecten in New Zealand described by Fleming (1957) with sharply defined to squarely margined radial ribs (Fig. 13A; colloquially known as the “square-ribbed” forms) appear at first glance to comprise a gradual evolutionary series. It commences with an extreme form, “P. tainui”, with high ribs, and seems to progress gradually through the less extreme “aotea form” with lower ribs, to evolve (apparently anagenetically) into the Recent P. novaezelandiae. The existence of this apparent evolutionary series may well have influenced Fleming’s thinking about the recognition of two “species groups” in New Zealand. Fleming (1957) also considered that there were two Recent geographical subspecies living in New Zealand, P. novaezelandiae novaezelandiae in the north and the more extremely “squareribbed” form P. novaezelandiae rakiura in the south. However, there are many complications in this scenario. Firstly, as the Recent “subspecies” have an unpredictable distribution, with the rakiura form (with the more sharply defined ribs) occurring in northern harbours as well as on open coasts in southern New Zealand, it is now generally agreed that the controls on this shell form are as much ecological as climatic, and the subspecies are not recognised in any recent reviews or catalogues of the New Zealand fauna. The variation in rib profile appears to be gradual (clinal) in any case. Woodburn’s (1990) demonstration of almost no electrophoretic distinction between the Nelson and Stewart Island populations also shows that the “subspecies” are trivial. Secondly, I collected a population from Landguard Sand (OIS 9), the type formation of P. novaezelandiae aotea, at Warrengate, Fordell, east of Wanganui (GS12779, S22/f67) of extremely “square-ribbed” specimens with high, undercut rib edges, more extreme than any seen from Tainui Shellbed at Castlecliff. Another specimen from Landguard Sand at Bruce Road, E side of Turakina valley (GS13567, S23/f04) has similar, undercut rib edges. It is clear, then, that the “aotea form” is not significantly different from the tainui phenotype of P. novaezelandiae, and the form is quite variable; no simple anagenetic series led up to the modern population of P. novaezelandiae. Also, other populations from high in the middle Pleistocene (at Te Piki, East Cape, and at localities around Wanganui) are intermediate in rib prominence between the “aotea” and “rakiura” forms. The strangest population is represented by an NMNZ collection from Rungapapa Knoll, W of White Island, Bay of Plenty, dredged in 188–228 m, recorded by Beu (2004, p. 155) under Crassostrea gigas (Thunberg, 1793). This collection was dated by 14C at 37 000–40 000 years, so they represent late Pleistocene fossils (OIS 3?) present in much deeper water than their living habitat, and so apparently deposited during a cool period of low sea level. In this population the radial ribs have markedly “squarecut” edges, and all the primary ribs are deeply subdivided into two; all specimens have twice as many primary radial ribs as in all other populations I have seen. The subtly different forms again are expressions of ecophenotypic rather than genetic variation. The marwicki phenotype (Fig. 13C; colloquially known as the “round-ribbed” form) differs from the tainui phenotype in its larger maximum size, its weaker inflation, the much lower primary radial ribs with a low, evenly convex cross section (mirror images of their interspaces) and obscurely defined edges, and the wider shape of large specimens, with concave dorsal outlines and low, weakly convex to weakly concave disc flanks extended anteriorly and posteriorly; that is, large specimens of this form develop a much wider umbonal angle than I have seen in the tainui phenotype. The larger size, wider flanks, and consequent wider umbonal angle might well result from the lesser expenditure on secreting tall-sided radial ribs than is necessary for the tainui phenotype. The tainui phenotype has much more clearly defined margins to the primary radial ribs, developed into vertical sides and a square-cut edge on extreme specimens (“P. tainui” and even more extreme forms, such as the population of “P. aotea” at Warrengate, east of Wanganui). The disc also remains evenly and relatively strongly convex and the outline evenly curved on the largest specimens of the tainui phenotype; no specimens have been seen that display the low, wide, anterior and posterior extensions of the disc flanks that occur in large specimens of the marwicki phenotype. Specimens from outcrops of Kupe Formation near the coastal exposures at Wanganui reach only a relatively small size and (presumably as a consequence) do not develop the widely extended disc flanks of large specimens, but remain evenly convex throughout life, and are subcircular in outline. They tend to develop a shallow median radial sulcus in the centre of each major radial rib. They therefore were thought by Fleming (1957) to be most closely related to the Western Australian form of Pecten fumatus, which has similarly sulcate primary radial ribs. As the Western Australian form was identified by Fleming (1957) by the name Pecten modestus Reeve, 1852 (now known to be a junior secondary homonym of P. modestus Bosc, 1802; = P. preissianus Iredale, 1949), he provided the name P. modestus kupei for the New Zealand fossil form (Fleming 1957, p. 37). The demonstration by Woodburn (1990) that the Western Australian form is little different from eastern Australian populations of P. fumatus (the slight electrophoretic difference presumably is explained by the gap of several thousand kilometres between her samples) indicates that the “kupei form” is not related to P. fumatus, and is part of the variation of the marwicki phenotype of P. novaezelandiae. This interpretation is confirmed by the geographical relationship of the “kupei form” to the marwicki phenotype in Kupe Formation, the only shellbed “Pecten modestus kupei” has been recorded from. Typical large, non-sulcate specimens of the marwicki phenotype occur at most localities more than a short distance inland from the coastal exposure. The “kupei form” is therefore interpreted here as based on a population of the marwicki phenotype that for some ecological reason remained at a small maximum size. Most localities where the “kupei form” occurs are near the seaward limit of outcrop, apparently the deepest deposition sites in the basin at that time that are now exposed on land, suggesting that such ecological factors as water depth or turbidity could explain the small shell size. Again, this is a purely ecophenotypic form, regarded here as a synonym of P. novaezelandiae. In my opinion, only one species of Pecten occurs in New Zealand, Pecten novaezelandiae Reeve, 1852.» BEU, A. G. 2006. Marine Mollusca of oxigen isotope stages of the last 2 million years in New Zealand. Par 2. Biostrattigraphically usefull and new Pliocene to Recent bivalves. Journal of the Royal Society of New Zealand, 36 (4): 151-338, figs. 1-41. [p. 205, 206-209]
|
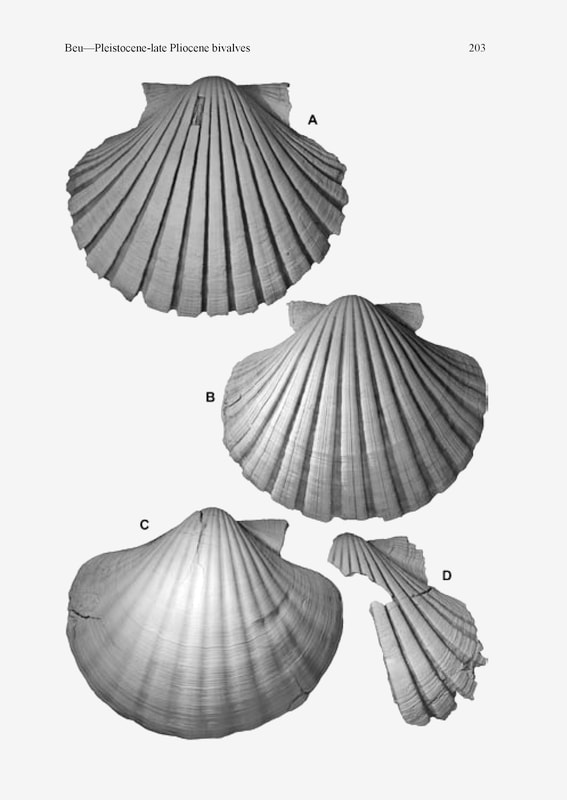
Pecten novaezelandiae (Reeve), all right valves from Wanganui Basin, ×0.66. A, tainui phenotype TM2015, GS4013, R22/f6353, Tainui Shellbed (OIS 13) at “the buttress”, Castlecliff (specimen illustrated by Fleming 1957, pl. 19, fig. 1), L 132.3 mm; B, specimen intermediate between tainui and marwicki phenotypes, holotype of Pecten jacobaeus toi Fleming, TM2027, GS4098, R22/f6425, uppermost Pinnacle Sand (OIS 14), “the pinnacles” gully, Castlecliff; L 123.4 mm; C,D, 2 phenotypes from one outcrop, GS13561, S22/f11, Pinnacle Sand (OIS 15/14), hillside above Kaukatea Valley Road, Okoia; (C), marwicki phenotype from fine sandstone, L 129.2 mm; (D), incomplete tainui phenotype from conglomerate lens, H 90.7 mm.; A. G. Beu, 2006, Marine Mollusca of oxigen isotope stages of the last 2 million years in New Zealand. Par 2. Biostrattigraphically usefull and new Pliocene to Recent bivalves, figure 13.
|
|
«Note on Pecten sulcicostatus 'var. casa van Bruggen, 1961'
This name, proposed (van Bruggen, 1961: 31, pl. 1, figs 1-3) as a infrasubspecific taxon, is not available, under ICZN Article 16. It was based on three specimens from the collection of an amateur, Mr John S. Hutt, made up of valves from six separate individuals. The material on which it was based is now in the Natal Museum (NMSA 87446, 87445), having been transferred there with the mollusc collection of the Albany Museum, Grahamstown, in which it was originally deposited. Although said by Hutt to have been trawled off Port Alfred, this material does not agree with anything known to occur in South Africa, but instead appears indistinguishable from Pecten novaezelandiae Reeve, 1852, from New Zealand.
The origin of this error is not difficult to deduce. Although Hutt specialised in collecting only Port Alfred shells, after his death in 1961 his collection proved to contain a substantial number of anomalous species, including such characteristic New Zealand taxa as Atalacmea fragilis (G. B. Sowerby lst, 1823), Trochus viridis Gmelin, 1791, and Buccinulum linea (Martyn, 1784). It cannot be coincidental that Hutt (pers. comm. to R. K., ca 1959) purchased many of his more unusual 'Port Alfred' specimens from a local shell-dealer!». KILBURN, R. N. & H. H. DIJKSTRA. 1995. A new species of Pecten Müller, 1776, from South Africa, with a note on "Pecten sulcicostatus var. casa" van Bruggen, 1961 (Mollusca: Bivalvia: Pectinidae). Annals of the Natal Museum, 36: 271-279, figs. 1-9. [p. 278, 279]
|

Pecten sulcicostatus Sowerby var. casa var. nov., holotype; A. C. van Bruggen, 1961, Notes on South African marine Mollusca. 4. A new variety of Pecten sulcicostatus, plate 1, figures 1-3.
|
   |