Kolymonectes weaveri Damborenea, 1998
DAMBORENEA, S. E. 1998. The bipolar bivalve Kolymonectes in South America and the diversity of Propeamussiidae in Mesozoic times. In P. A. Johnston & J. W. Haggard (Eds), 1998: An Eon of Evolution - Paleobiological studies Honoring Norman D. Newell, 143-155. University of Calgary Press. Calgary. [p. 150, pl. 1, figs. 1-16, text-figs. 1A, 1B, 2A-2C]
1931 Pecten (Variamussium) personatus var. coloradoensis Weaver, 1931
1998 Kolymonectes weaveri Damborenea, 1998
1998 Kolymonectes weaveri Damborenea, 1998

S. E. Damborenea, 1998, plate 1.


S. E. Damborenea, 1998, figures 1, 2.
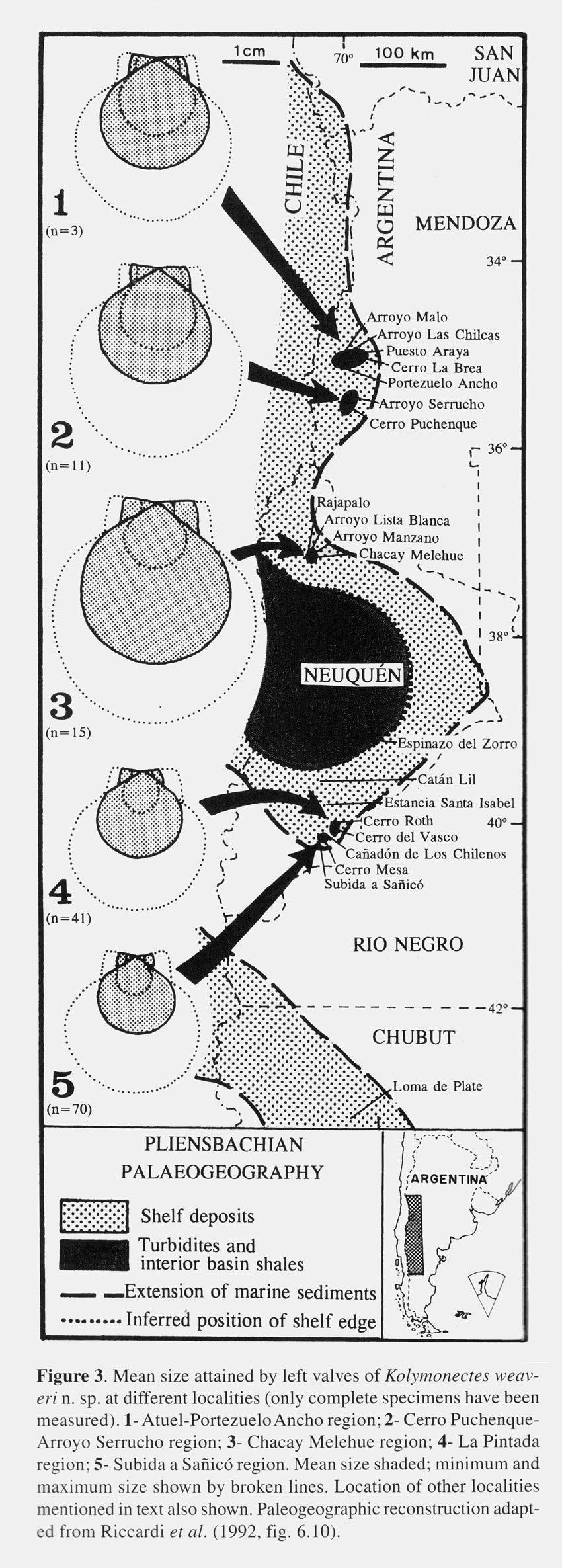
S. E. Damborenea, 1998, figure 3.
|
«Kolymonectes weaveri n. sp.
PI. I, Figs. 1-16; Figs. 1 (A-B), Figs. 2 (A-C)
Etymology. After Charles Weaver who first recognized this as a different taxon in his outstanding monograph on Mesozoic Argentinian molluscs.
Material. Holotype-MLP 23686, a complete specimen with valves slightly displaced, preserved as internal and external moulds, from between Cerro Roth and Cerro Del Vasco, Piedra Pintada region, southern Neuquen (PI. I, Figs. 1-3).
Other material—about seven hundred specimens, most of them preserved as external or internal moulds of isolated valves. A few specimens have fragments of shell material and about a dozen have both valves preserved, though not usually articulated (MLP 5809, 6000-6012, 6017, 6031, 6102, 6209, 6667, 6675, 6686, 14407, 14415, 16189, 16194, 16425, 18203, 19675, 23685, 23687-23742, 23744-23783, 23785-23801, 23803-23814, 26560, 27549-27555). Also included-holotype (BMW SA 15511029 [= cast MLP 22355]) of Pecten coloradoensis Weaver: external mould of a left valve, almost complete but damaged during preparation, from Weaver's locality 1029, immediately to the north of Catan Lil, Neuquen (Weaver, 1931, pI. 28, fig. 164 and PI. I, Fig. 5 herein). Weaver (1931, p. 275) erroneously mentioned that the specimen was found in Mendoza. Diagnosis. Orbicular, inequivalve, and slightly inequilateral shells. Posterior and anterior auricles almost equal, separated from disc by deep sutures. Shallow byssal notch without ctenolium. Disc orbicular with flatter anterior and posterior flanks. Umbonal angle wider than 100°. Right valves ornamented by very faint radial lines; interior smooth. Left valves with narrow radial plicae separated by wide intervals and closely set comarginal growth-lines. Radial plicae increase in number by intercalation. Outer surface of auricles smooth except for faint radial lines on left posterior auricle. Subtriangular, centrally placed resilifer with one elevated ridge to each side in the right valve. Each auricle with one long cardinal crus.
Description. Shells medium-sized, orbicular, inequivalve, and slightly inequilateral. Shell convexity low and nearly equal on both valves. Young shells higher than long, differences in these measurements diminishing with size. Some large valves longer than high. Young shell orthocline, becoming somewhat prosocline with growth, with posterior portion larger than anterior (PI. I, Fig. 16). Anterior and posterior auricles subequally long. Right auricles externally convex; left auricles concave. Right auricles neatly separated from disc by deep suture; left valve sutures less deep though still clear. Right
anterior auricle with very shallow byssal notch lacking ctenolium (PI. I, Figs. 8, 10-11). Left anterior auricle sometimes showing traces of byssal sinus. Umbonal angle always wider than 100°, tending to increase with size. Umbones low and inconspicuous. Dorsal margin slightly less than half total shell length. Anterior auricles with dorsal margin on crest of low fold increasing in amplitude distally and bearing short projecting spines. Angle of dorsal margins of right valve about 170°. Anterior and posterior auricle margins meeting dorsal line at an obtuse angle; posterior auricle margin less obtuse. Anterior and posterior margins of disc straight but evenly merging into rounded ventral margin. Angulation variably present on right valves where posterior and ventral margins of disc meet (PI. I, Figs. 1-4, 8, 10), these valves having lost outermost shell layer. 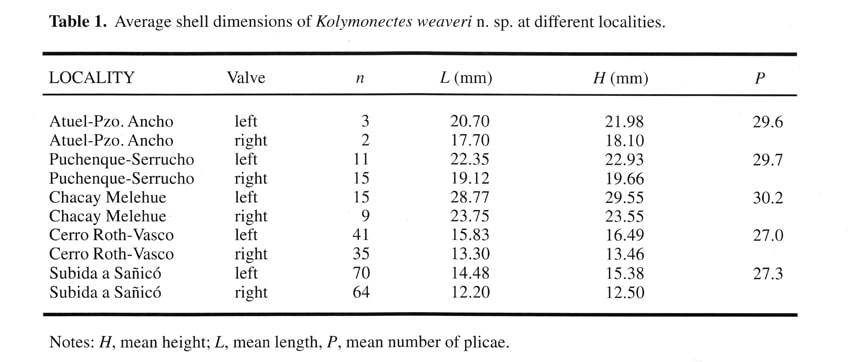
Anterior and posterior regions of disc flatter than central portion, especially on posterior region of left val ve which has abrupt change in shell convexity.
Ornament pattern differing on each valve (Figs. 2A, 2B). Right valves covered sub-externally by very faint closely set radial lines (PI. I, Figs. 8, 12), and internally smooth except sometimes for narrow marginal band (Fig. 1B). Left valves ornamented by up to 40 (mean 29) narrow radial plicae separated by wide intervals (PI. I, Figs. 1-3, 7-8) and delicate and closely set comarginal growth lines (Fig. 2A). Radial plicae increase in number by intercalation in somewhat irregular pattern. Primary plicae evident on internal views (PI. I, Fig. 6). Growth lines strongest on tops of plicae. Radial plicae straight on anterior half ofleft valve but tending to curve backwards on posterior region of disc. Left posterior auricle bearing few, faint, radial lines (PI. I, Figs. 7, 8); other auricles smooth with regular growth-lines only. One long cardinal crus extending to each side of sub-triangular and centrally placed resilifer. Resilifer of right valve laterally limited by one short ridge on each side. Right valve sutures internally thickened but not forming true auricular crura. Other internal characters unknown. Shell very thin. Shell dimensions vary according to locality (Table 1 and Fig. 3). Remarks. This species is the most abundant pectinoidean in Pliensbachian deposits of Argentina. It includes the specimen described by Weaver (1931) as Pecten (Variamussium) personatus var. coloradoensis. The name Pecten (Monotis?) coloradoensis had been given to a Permian species from North America (Newberry, 1861) and later merely listed under Aviculopecten (Girty, 1903, p. 48). It was then transferred to Acanthopecten (Newell, 1937;. Newell and Boyd, 1995). Judging from ICZN articles 52b, 53c and 57b, Newberry's and Weaver's species-group names are thus primary homonyms. Hence, a valid name is lacking for the latter. In this particular case, straightforward proposal of a replacement name is not advisable in view of the inadequate preservation of Weaver's type specimen (PI. I, Fig. 5) as well as uncertainties about its geographic and stratigraphic provenance. Therefore, a new species is here erected for this important taxon, with Weaver's name thus falling into subjective rather than objective synonymy. This is supported by a suitable holotype from an accessible type locality and well-defined type horizon where the species is known to be frequent. Despite its great abundance, this species was poorly known previously. Furthermore, both Weaver (1931) and Leanza (1942) assumed that the radial external ornament present on the left valve represented internal ribs. As a consequence, they referred this species to Variamussium and their descriptions were not only incomplete but misleading. Weaver (1931) apparently had only a very poorly preserved left valve, yet he stated that the shell was inequivalve. Leanza had more than a hundred specimens but failed to notice that the ornament was different on each valve and simply declared that the exterior of the shell was smooth and the interior ribbed. He even figured (1942, pI. 10, fig. 3) a composite mould of a left valve as an "inner surface of right valve". Though realizing that the costae were external, Johnson (1984, p. 28) doubtfully referred the material figured by Leanza (1942) to Propeamussium (P.) laeviradiatum (Waagen).
At least some of the specimens described by Weaver (1931) as "Oxytoma inaequivalve" (Sowerby) from Piedra Pintada may also belong here. His description and especially his figure (pI.19, fig. 87) agree with left valves of K. weaveri, this species also being very common at the locality mentioned by Weaver. Affinities. Kolymonectes anjuensis (Milova) from the Sinemurian of northern Russia (Milova, 1969, pI. 1, figs. 1-9; 1976, pI. 11, figs. 1-9) is very similar to K. weaveri in outline and ornament, but the type species of Kolymonectes has a much smaller shell and relatively more radial plicae. Of all boreal species, K. staeschei (Polubotko) is most similar to the Argentinian species and is of late Sinemurian-early Pliensbachian age in northeast Russia and Hettangian-Sinemurian in Canada (Polubotko, 1968, pI. 7, figs. 1-9; Polubotko and Milova, 1986, pI. 15, figs. 9-13; Milova, 1988, pI. 5, figs. 9-27; Poulton, 1991, pI. 6, figs. 21-28; Sey and Polubotko, 1992, pI. 120, figs. 26-27). Right valves of this species show a marginal band of radial plicae which could be a reflection of the inner margin of left valves through the thin and flexible marginal apron of the prismatic shell layer. Radial plicae on left valves are more numerous and irregular than in K. weaveri and have beaded tops.
Kolymonectes kedonensis Polubotko (in Polubotko and Milova 1986, pI. 15, figs. 1-8), from the Hettangian (?) Sinemurian of northeastern Russia, has fewer radial ribs with strongly beaded tops. From the same region, the Pliensbachian species K. monkgensis Polubotko (in Polubotko and Milova 1986, pI. 15, figs. 14-19) has similar radial ornament but this is superimposed by concentric irregular folds which are absent in K. weaveri. Kolymonectes terekhovi (Polubotko, 1968, pI. 26, figs. 2-3; Milova, 1988, pI. 6, figs. 13-15) from the Pliensbachian of the Alazei Plateau has strong internal ribs that limit the disc flanks of the right valves; these are absent in K. weaveri. Compared with species referred to Ochotochlamys Milova and Polubotko by Russian authors, K. weaveri differs mainly in having a very shallow byssal notch. "Pecten" newberryi Whitfield (1880, pI. 4, figs. 12-15), from the Jurassic beds of the Black Hills of North Dakota, is comparable to K. weaveri in shape and ornament, but radial ornament on the left valve is made up of more numerous fine costae which are hardly reflected on the inner surface of the shell. The right valve figured by Whitfield was not available with the other syntypes (AMNH CU 9562 G). Unfortunately, no accurate data regarding the age of this interesting species are available, and more material is needed to ascertain its relationships with Kolymonectes. Some coarsely ribbed specimens of Jurassic to Tertiary Variamussium species have internal radial ribs but show a remarkable resemblance to K. weaveri in external appearance. These include the left valves figured by Holder (1978, pI. 1, figs. 1-2) as Parvamussium pumilus atlasense Holder; by Polubotko (1968, pI. 66, figs. la-Ib) as Variamussium sp.; by Moore (1984, pI. 2, figs. 3, 5, 7) as Parvamussium alaskaensis (Dall); by Johnson (1984, pI. 1, figs. 5-6) as Propeamussium laeviradiatum Waagen; and by Damborenea (1992, pI. 118, fig. 5) as P. andium (Tornquist). Occurrences. Kolymonectes weaveri occurs from southern Mendoza to Chubut, at the following localities, from north to south (Fig. 3; for precise locations see Damborenea, 1987a, 1987b): Puesto Araya and Cerro La Brea, Arroyo Serrucho, Cerro Puchenque (Mendoza Province); AlToyo Lista Blanca, Cordillera del Viento, Arroyo Chacay Melehue, Arroyo Manzano, Espinazo del Zorro, South of Estancia Santa Isabel, NW of Puesto Manqueu, Subida a Sañicó, SW of School 27, Cerro Mesa, Cerro Del Vasco, Cañadón La Pintada, between Cerro Roth and Del Vasco, Cerro Roth, Cañadón de Los Chilenos (Neuquén Province); Loma de Plate (Chubut Province).
This species ranges in age from late Early Pliensbachian to latest Pliensbachian, i.e., it mostly coincides with the Fanninoceras Ammonite Zone in the sense of Riccardi (1984) and has been found both in the Otapiria neuquensis and Radulonectites sosneadoensis Assemblage Zones based on bivalves (Damborenea in Riccardi et al., 1990).» SUSANA ESTER DAMBORENEA, 1998
|