Cryptopecten nux nux (Reeve, 1853)
REEVE, L. A. 1855. Conchologia Iconica: or Illustrations of the Shells of Molluscous Animals , vol 8. London, 1855. 35 pls [pls 1-12 + text (1852); pls 13-35 + text + index (1853)]. Unpaginated [pl. 32, fig. 143 (coruscans in plate text, description in next plate, sp. 149)]
1853 Pecten nux Reeve, 1853
1888 Pecten guendolenae Melvill, 1888
1908 Chlamys smithi G. B. Sowerby III, 1908
1909 Chlamys corymbiatus Hedley, 1909
1934 Pecten (Aequipecten) kikaiensis Nomura & Zinbo, 1934
1888 Pecten guendolenae Melvill, 1888
1908 Chlamys smithi G. B. Sowerby III, 1908
1909 Chlamys corymbiatus Hedley, 1909
1934 Pecten (Aequipecten) kikaiensis Nomura & Zinbo, 1934

L. A. Reeve, 1853, plate 32.
|
«PECTEN CORUSCANS. Pect testa globosa, valde convexa, aquilaterali, inaequivalvi, valvis costis peculariter acute tri-liratis vigintl radiatis, interstitiis dense scabroso-cancellatis; albida, rosea varie tincta et maculata; auriculis inaequalibus.
THE GLITTERING PECTEN. Shell globose, very convex, equilateral, inequivalve, valves rayed with twenty ribs, which are peculiarly sharply three-ridged, interstices densely scabrous-cancellated; whitish, variously stained and blotched with rose; ears unequal. HINDS, Moll. Voy. Sulphur, p. 61. pl. 17. f. 3. Hab. Port Anna Maria, Nukuhiva, Marquesas Islands (in seven fathoms on a sandy floor); Belcher. The ribs of this species differ peculiarly in structure from those of any other.» LOVELL AUGUSTUS REEVE, 1855
|
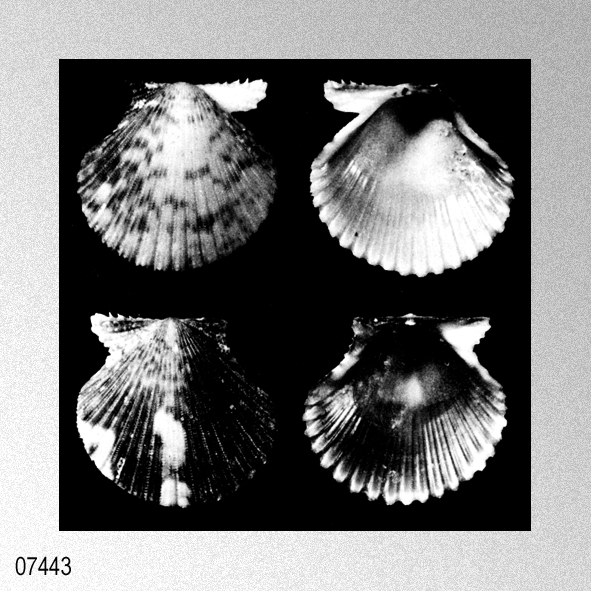
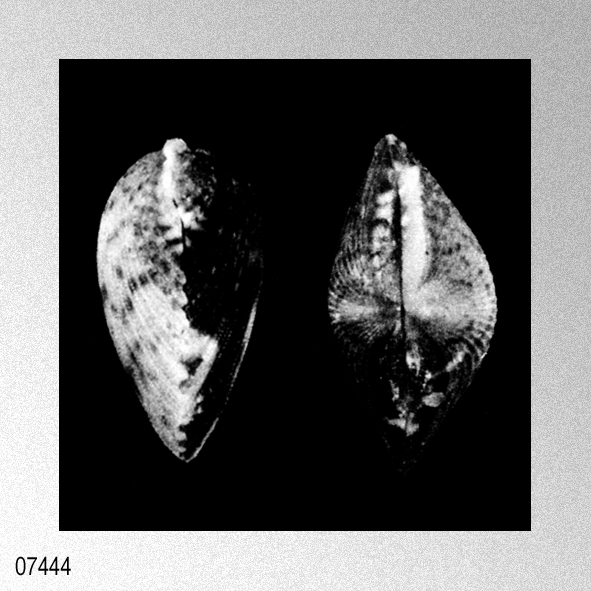




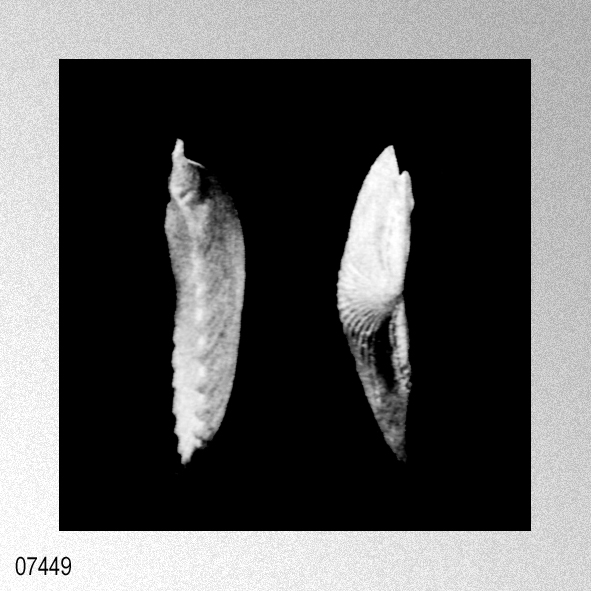














|
«Distribution: Throughout tropical and subtropical Indo-West Pacific (absent from Hawaii), south-western limit western Transkei.
Habitat: Living byssally attached to rocks or coral, or on sediments or coral rubble on sandy bottoms at littoral to bathyal depths. Remarks: C. nux is rather variable in outline, convexity, sculpture and colour (Hayami 1984:100; Dijkstra 1991: 37). Type material of P. guendolenae, C. smithi and C. corymbiata is morphologically similar to C. nux. Wagner (1989: 58) considered C. guendolenae to be a valid species, but Hayami (1984: 102) and Dijkstra (1991: 37) regarded it as a synonym of C. nux. C. bernardi (Philippi, 1851) from French Polynesia is larger in size (up to 25 mm high), more triangular in shape (when mature) and more convex, and has finer sculpture than C. nux. However, it is possible that the type material of C. nux consists of young specimens of C. bernardi. Similar material has been observed from French Polynesia (HD, MNHN, NNM, ZMA) (under study). South African material, misidentified by Barnard (1964: 429) as ‘Chlamys bullatus Dautzenberg & Bavay’, is in all features similar to C. nux.» DIJKSTRA, H. H. & R. N. KILBURN. 2001. The family Pectinidae in South Africa and Mozambique (Mollusca: Bivalvia: Pectinoidea). African Invertebrates, 42: 263-321. [p. 312]
|

Cryptopecten nux (Reeve, 1853); H. H. Dijkstra & R. N. Kilburn, 2001, The family Pectinidae in South Africa and Mozambique, figures 50, 51.
|
|
«Distribution: Red Sea, east coast of Africa to South Africa, Réunion, Mauritius, Arabian Gulf, extending eastwards to southern Japan and New Caledonia. Littoral to bathyal depths, usually sublittoral, living among coral rubble on soft sediments (sand or muddy sand). For more detailed information on distribution and geographical variation see Hayami (1984: 99, 102).
Remarks: Specimens of C. nux from the Red Sea are identical in morphological characters to the type specimens. For description of the present species see Hayami (1984: 101) and Dijkstra (1991b: 37). For more information and a comparison with related species see Hayami (1984: 102).» DIJKSTRA, H. H. & J. KNUDSEN. 1998. Some Pectinoidea (Mollusca: Bivalvia: Propeamussiidae, Pectinidae) of the Red Sea. Molluscan Research, 19 (2): 43-104, pls. 1-10. [p. 87]
|

Cryptopecten nux (Reeve, 1853); H. H. Dijkstra & J. Knudsen, 1998, Some Pectinoidea of the Red Sea, plate 3, figures 12, 13.
|
|
«Remarks. — Originally I considered C. nux a synonym of C. bernardi, a point of view that was generally shared by many others. As explained under C. bernardi, this initially seemed justified, especially in the light of the poor knowledge of the group. Another species easily mixed up with C. nux is C. guendolenae (Melvill, 1888). A comparison of the relative ratio's of the complete shells and the right valves of C. nux and C. guendolenae, shows clearly that C. nux has a more convex right valve as well as a more globose shell in general than C. guendolenae (figs. 17-18). Due to the fact that some species of Cryptopecten occur sympatrically, and that I only examined the figured syntype of C. nux, I do not know whether the other syntype material belongs to the same species only, and therefore I consider it appropriate to designate the figured syntype to be lectotype of C. nux.
Although the condition of the South African material examined is rather poor, it can be assigned to this species on the basis of the globose right valve, smooth lunulae, and, above all, the number of ribs, that constantly differs from that of C. guendolenae. As explained under C. bernardi the original type-locality is most probably wrong, and in order to avoid confusion in future it seems justified to designate a new typelocality for the species. Type-locality (new designation). — Panglao, Bohol, Philipines Distribution.— Indian Ocean and Western Pacific.» WAGNER, H. P. 1989. The genus Cryptopecten Dall, Bartsch & Rehder, 1938, in the Indo-Pacific (Mollusca; Bivalvia; Pectinidae). Basteria, 53: 53-62, figs. 1-18. [p. 57, 58]
|

H. P. Wagner, 1989, The genus Cryptopecten Dall, Bartsch & Rehder, 1938, in the Indo-Pacific. C. nux (Reeve), figures 6-9; C. guendolenae (Melvill), figures 10-13.
|
|
«Diagnosis.— Tumid, smallest species of Cryptopecten characterized by relatively strong inflation of both valves, large anterior wing, 18-22 apparently tripartite radial ribs, regularly alternate fine imbricated scales on radial ribs and moderately thick test.
Remarks.— C. nux is similar to C. bullatus and different from C. vesiculosus in the small shell size, large number of radial ribs, slender central ridge of each rib, and regularly alternate disposition of imbricated scales. However, the convexity of both valves is generally much stronger in comparison with the other two species (see Figs. 9, 10). The inner shell layer is poorly developed in C. bullatus but attains considerable thickness in the present species as well as in C. vesiculosus. Dr. T. R. Waller suggested to me (oral communication) the possibility that Pecten nux Reeve, 1853, together with several subsequently proposed nominal species, may fall into a junior synonym of Pecten bernardi Philippi, 1851, because one of the "type-specimens" of. P. bernardi with no locality data, which is preserved in the British Museum (Natural History) (BM (NH) 1923.7.13.7), shares essentially the same morphological characters with the illustrated syntype of P. nux. As he commented, however, some ambiguity still remains as to the type identity, because Philippi's original description was brief and not accompanied by any illustrations. For the time being I regard P. nux as the earliest undoubted name for the species under consideration. Among several fossil samples of this species from Japan, the Late Pleistocene sample Sm (N) shows peculiar morphological features deserving a subspecific distinction. Consequently, all the other fossil and Recent samples treated here are provisionally assigned to the nominate subspecies, Cryptopecten nux nux. Geographic variation.— In addition to several fossil and Recent samples from central and south Japan, I have examined numerous well-documented Recent samples of this species from extensive areas of the Indo-Pacific which are preserved in the National Museum of Natural History, Washington D. C. These samples often differ from one another in such characters as shell size, convexity, obliquity, thickness of test, and shape and sculpture of byssal wing. The difference must be partly due to the age heterogeneity but is mostly attributable to geographic variation. The average number and tripartite appearance of radial ribs, however, hardly vary at all. It is my impression that the center of distribution of this species is in the Philippine Islands, though it seems to be also common in some areas of Melanesia, Polynesia, Queensland and the Indian Ocean. Most of the specimens from the Philippines are characterized by relatively small and right-convex shell and moderate thickness of test. The fossil and Recent samples from various localities in south Japan are not much different from these specimens in shell morphology. On the other hand, the observed specimens from the Marquesas Islands, the type locality of P. nux, as well as those from New Britain, show thicker test and more equiconvex shell. The material from Queensland, as exemplified by the specimen USNM 764157 (Plate 9, Fig. 2) and the illustrated syntype of Chlamys corymbiatus, possesses thick test and occasionally weak tubercles on the central solid ridge of each radial rib, which are seemingly absent in the specimens from other regions. Generally speaking, the specimens from the Indian Ocean appear to be more weakly inflated than those from the Philippines and many other areas of the Pacific. The holotype of Chlamys smithi Sowerby, 1908, from Mauritius Island, seems to exemplify the Indian Ocean form. Moreover, the examined sample (USNM 764165) from the northern part of the Red Sea (eastern coast of Egypt) is unique for the nearly equiconvex and unusually weakly inflated valves (Plate 9, Fig. 5). The geographic variation of C. nux is thus considerably wide. If subspecific division were applied to this species, the trivial names corymbiatus and smithi (or guendolenae) would be available for the populations of Queensland and the main part of the Indian ocean, respectively. The morphological change in these areas, however, may be gradational, and all the observed samples except a few Philippine ones are too small in sample size to recognize clines or to detect significant morphological difference between local populations. Cryptopecten nux nux (Reeve)
Pl. 2, Fig. 4; Pl. 3, Figs. 1, 2; Pl. 9, Figs.2-5; Pl.12, Figs. 1, 2 1853. Pecten coruscans Hinds: Reeve, Conchologia lconica, vol. 8, pl.32, fig. 143. [non Pecten coruscans Hinds, 1845]
1853. Pecten nux Reeve, Conchologia lconica, vol. 8, errata. 1888. Pecten hastingsii Melvill, Jour. Conchology, vol. 5, p. 279, pl. 2, fig. 7 . 1888. Pecten guendolenae Melvill, Jour. Conchology, vol. 5, p. 279, pl. 2, fig. 6. 1888. Pecten nux Reeve: Küster and Kobelt in Martini and Chemnitz, System. Conch. Cab., vol. 7, pt. 2, p.163, pl. 45, figs. 5-8. 1908. Chlamys smithi Sowerby, Proc. Malacol. Soc. London, vol.8, p. 18, pl. 1, figs. 6, 7. 1909. Chlamys corymbiatus Hedley, Proc. Linn. Soc. New South Wales, vol. 34, p. 423, pl. 36, figs. 1-4. 1912. Pecten (Aequipecten) vesiculosus Dunker: Dautzenberg and Bavay, Les Lamell. Expéd. Siboga, System., 1. Pectinidés, p. 148. [non Pecten vesiculosus Dunker, 1877] 1930. Chlamys (Aequipecten) nux (Reeve): Cox, Monogr. Geol. Dept. Hunterian Mus., Glasgow Univ., vol. 4, p. 124, pl. 14, fig. 11. 1933. Pecten (Aequipecten) nux Reeve: Nomura, Sci. Rep. Tohoku Imp. Univ., ser. 2, vol. 16, no. 1, p. 55. 1934. Pecten (Aequipecten) nux Reeve: Nomura and Zinbo, Sci. Rep. Tohoku Imp. Univ., ser. 2, vol. 16, no.2, p. 117 (pars). 1934. Pecten (Aequipecten) kikaiensis Nomura and Zinbo, Sci. Rep. Tohoku Imp. Univ., ser. 2, vol. 16, no. 2, p. 153, pl. 5, fig. 9a, b. 1939. Corymbichlamys corymbiatus (Hedley): Iredale, Brit. Mus. (Nat. Hist.) Sci. Rep., vol. 5 (Mollusca pt. 1), p. 368. 1951. Cryptopecten nux (Reeve): Habe, Genera of Japanese Shells, p. 77. 1956. Chlamys (Aequipecten) nux (Reeve): Eames and Cox, Proc. Malacol. Soc. London, vol.32, p. 43. 1961. Cryptopecten nux (Reeve): Habe, Coloured Illustrations of the Shells of Japan (II), p. 118, pl. 53, fig. 9. 1964. Chlamys (Cryptopecten) nux (Reeve): Shikama, Selected Shells of the World II, p. 50, fig. 93. 1964. Cryptopecten nux (Reeve): Habe, Shells of the Western Pacific in Color, vol.2,p. 174, pl. 53, fig. 9. 1977. Cryptopecten nux (Reeve): Habe, Systematics of Mollusca in Japan, p. 84. 1977. Cryptopecten kikaiensis (Nomura and Zinbo): Habe, Systematics of Mollusca in Japan, p. 84. 1977. Cryptopecten hastingsii (Melvill): Habe, Systematics of Mollusca in Japan, p. 85. 1982. Cryptopecten nux (Reeve): Hayami, Venus, vol. 47, p. 235 Type.— Reeve's illustrated syntype (BM (NH) 1950. 11.14.52), which was erroneously named "Pecten coruscans" in the original text but subsequently corrected in the index and errata, is a conjoined specimen from Marquesas Island of Polynesia. It is preserved in the British Museum (Natural History). ca. 15 mm long, 14 mm high, 8 mm thick. The holotype of Pecten hastingsii from Japan (ca.25 mm long, 24 mm high, 14 mm thick) and the holotype of Pecten guendolenae from Mauritius (ca. 15 mm long, 14 mm high, 7 mm thick) are conjoined specimens preserved in the Cardiff Museum (Tomlin Collection). The holotype of Chlamys smithi from Mauritius (BM (NH) 1908.5.30.63) (ca. 16 mm long, 16 mm high, 7 mm thick) is a conjoined specimen preserved in the British Museum (Natural History). The syntypes of Chlamys corymbiatus are said to have been presented to the Australian Museum. The holotype of Pecten (Aequipecten) kikaiensis is a right valve (IGPS no. 50357) from the Late Pleistocene Wan Formation at Kamikatetsu of Kikai Island, south Japan, which is preserved in the Institute of Geology and Palaeontology, Tohoku University.
Material.— specimens used for the following description and discussions are indicated in the list of examined samples (p. 123). Diagnosis.— Subspecies of C. nux, possessing variably but usually strongly inflated valves and narrowly elongated byssal wing with prominent radial ribs and often with spiny scales. Description.— Shell small, rarely exceeding 20 mm in length and height, highly inequivalve, nearly acline or very slightly prosocline, more or less right-convex except for very early growth stages; height negatively allometric to length, commonly subequal to length in young stages but becoming considerably smaller than length with growth; thickness nearly isometric to length in each valve; form ratio T/L greatly variable even within a sample, ranging 0.23-0.37 in adult right valves and 0.17-0.26 in adult left valves; hinge line moderate or comparatively long, negatively allometric to overall length; test moderate but somewhat variable in thickness; apical angle of disk 100-105 degrees; anterior wing more than twice as long as posterior; byssal wing generally slender, with elevated dorsal margin above hinge axis; anterodorsal and posterodorsal margins of disk nearly straight, subequal in length, scarcely gaped between valves; byssal notch moderate in depth, with four or five exposed denticles of ctenolium and fasciole area of moderate width. Outer surface of disk ornamented with 18-22 radial ribs, each of which consists of a solid, roof-shaped central ridge and a pair of hollow parts on its sides covered by fine alternate imbricated scales; accordingly, if scales are exfoliated or removed, radial ribs look regularly tripartite; imbricated scales numbcring eight to nine per 1 mm on middle-ventral surface of adult shell; interspace of ribs as wide as rib, marked with short erect scales, which are independent from and generally more distantly spaced than imbricated scales on ribs; intercostal radial threads commonly absent but rarely observed in some adult individuals; growth ring(s), if present, very weak; disk sculpture of bothvalves essentially similar; byssal wing generally possesses four strong radial ribs on which several spiny scales frequently occur; other wings also ornamented with a few radial riblets. Coloration variable, sometimes monotonously yellow but more commonly reddish brown in ground color with irregular pale bands and spots; left valve generally more darkly pigmented than right, not only externally but also internally; byssal wing always pale in color. Internal surface somewhat shiny, strongly plicated in accordance with radial ribs, but umbonal to middle part much flattened by deposition of inner shell layer; musculature weakly impressed; resilial insertion moderate in size; cardinal crura developed but not very strong. Remarks.— This subspecies seems to include almost all the specimens hitherto described under the names of Pecten nux and Chlamys corymbiatus and also the type materials of Pecten hastingsii, Pecten guendolenae, Chlamys smithi and Pecten (Aequipecten) kikaiensis. In spite of an extensive geographic distribution in the Indo-Pacific, the present subspecies seems to be comparatively rare in the seas around the Japanese Islands: every Recent sample consists of one or a few individuals. Only the sample Kk (N) from the Late Pleistocene of Kikai Island is composed of a large number of individuals, allowing quantitative examination of intrapopulational variation and relative growth (Tables 3, 4, 6-9). At the same locality two other species of Cryptopecten, i. e., C. bullatus and C. spinosus, occur in association, and they are clearly distinguishable from the present subspecies. As suggested from this sample as well as such samples as Bh (N), USNM 230228, 230314 and 292450 from the Philippines, the intrapopulational variation of shell outline and convexity is considerably wide; the right valve is generally characterized by strong convexity, but the form ratio T/L is quite variable, as indicated by the relatively low value of correlation coefficient between log L and log T (or log C). Besides the sample Kk (N), C. nux nux is represented by several small samples of various geological ages from the Ryukyu Islands (samples Mk (N), Ik (N) and Ob (N)). Though their morphometric comparison is still difficult owing to their small sample size, essential morphological characters including the mode and number of radial ribs appear almost unchanged throughout the geological period. Neogene fossils of C. nux are are considerably well represented in the tropical coastal region of east Africa. Cox (1930) and Eames and Cox (1956) recorded occurrences of this species from the Lower Miocene at several localities in Zanzibar and Pemba Islands in Tanzania, and also from the Pliocene near Mombasa in Kenya. Although I have had no opportunity to observe the African specimens, it is quite likely that the range of ihis species extends down into the Early Miocene. For the time being I regard these fossils as belonging to C. nux nux. Distribution.— This subspecies is widely distributed in the tropical and subtropical seas of the central-west Pacific and the Indian Ocean. Although dead shells have been dredged from outer sublittoral and bathyal bottoms, the hitherto known records of living shells are almost entirely confined to the inner sublittoral zone. Recent specimens have been known from Polynesia (especially Marquesas and Tahiti), Melanesia, Micronesia, northeast Australia, Indonesia, Singapore, the Philippines, the South China Sea, Formosa, south Japan, Bengal Bay, Andaman, Oman, Seychelles, Mauritius, Madagascar, Mozambique, east coast of South Africa, and the Red Sea. There is no undoubted record of occurrence in the Japan Sea. Fossils of this subspecies occur from the Lower Miocene of east Tanzania (Eames and Cox, 1956), the Pliocene of east Kenya (Cox, 1930), and the Pliocene Byoritsu Formation of Formosa (Nomura, 1933) in addition to the Pliocene and Pleistocene in central and south Japan (see the list of examined samples). Early Miocene to Recent.» HAYAMI, I. 1984. Natural history and evolution of Cryptopecten (A Cenozoic-Recent Pectinid Genus). The University Museum, The University of Tokyo, Bulletin, 24: 1-149, pls. 1-13. [p. 99-103]
|



Cryptopecten nux nux (Reeve); I. Hayami, 1984, Natural history and evolution of Cryptopecten, plate 2, figure 4 (above); plate 3, figures 1, 2 (middle); plate 9, figures 2-5. (below).
|
|
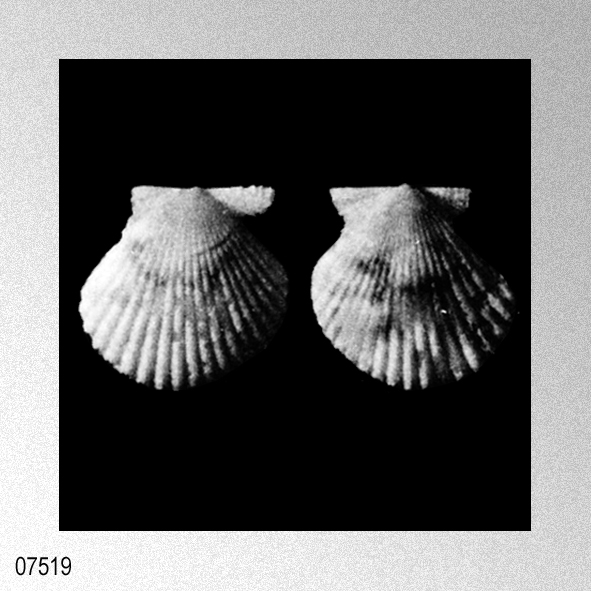
«1930. Chlamys (Aequipecten) nux (Reeve), Cox, (15), p. 124, PI. 14, fig. 11. (cum syn.).
Material:— Sample APT.82 (Zanzibar, spoil from well N. of road at Kwebona; L. Miocene, with Lepidocyclina), 3 specimens; sample APT.133 (Zanzibar, from 12 ft. cliff near Shangani; L. Miocene), 1 specimen; sample APT.134 (Zanzibar, 700 yds. N. along coast from Shangani; L. Miocene), 1 specimen; sample APT.256 (Pemba 1., 20 ft. cliff on S.W. coast of Pakaima peninsula, 1,900 yds. at 324° from Ras Domoni; L. Miocene), 1 specimen; sample REL.3006 (Kenya, Mtwapa Creek, N. side, 1½ miles up from ferry; Pliocene), 1 specimen.
Remarks:— Quoted in the paper cited as occurring in the Recent faunas of Mauritius, Bay of Bengal, Siam, Sulu Archipelago, Tahiti, Marquesas Is., Gulf of Oman, and Maldives, and recorded also from the Pliocene near Mombasa. The specimens from APT.82, which are associated with Lepidocyclina, confirm the extension of the range down into the L. Miocene.» EAMES, F. E. & L. R. COX. 1956. Some Tertiary Pectinacea from East Africa, Persia, and the Mediterranean region. Proceedings of the Malacological Society of London, 32 (1): 1-68, pls. 1-20. [p. 43]
|
|
«Localities. Nos. 4 [Cliff, north-east of Mombasa, south of the Ras Makawe Jini] and 6a [Cliff, north-west of Mombasa, at Schimanzi] (several specimens).
Further Distribution. Fossil: No previous records. Recent : Mauritius, Bay of Bengal, Siam, Sulu Archipelago, Tahiti, Marquesas Is. (Dautzenberg and Bavay), Gulf of Oman (Melvill), Maldives (Smith). Remarks. This is a small, orbicular, gibbose, sub-equivalve species, ornamented with about twenty not very prominent, but angular ribs, which are separated by slightly narrower interspaces. The finer details of the ornamentation are very characteristic, the fossils agreeing perfectly with Recent specimens. Each rib bears three sharp longitudinal ridges, one at its crest, and one on each side. Between the lateral ridges there extend delicate, transverse, chevron-shaped lamellae, the apex of the chevron lying on the median ridge. The intercostal spaces are ornamented with regularly spaced transverse lamellae, which are slightly more prominent and further apart than the chevrons. The anterior auricle of the right valve is very narrow, with its lower margin parallel to the hinge-margin and the byssal notch deep» COX, L. R. 1930. Reports on Geological Collections from the Coastlands of Kenya Colony made by Miss M. McKinnon Wood. V, VI, VII : Miocene, Pliocene, Post-Pliocene Mollusca. Monographs of the Geological Department of the Hunterian Museum, Glasgow University, 4: 103-163, pls. 12-15. [p. 124]
|

Chlamys (Aequipecten) nux (Reeve); L. R. Cox, 1930, Reports on Geological Collections from the Coastlands of Kenya Colony made by Miss M. McKinnon Wood, plate 14, figure 11.
|