Cryptopecten vesiculosus (Dunker, 1877)
DUNKER, W. 1877. Mollusca nonnulla nova maris japonici. Malakozoologische Blätter, 24: 67-75. [p. 72]
1877 Pecten vesiculosus Dunker, 1877
1877 Pecten trifidus Dunker, 1877
1882 Pecten jickelii Dunker, 1882 [nomen novum pro Pecten trifidus Dunker, 1877]
1888 Pecten hysginoides Melvill, 1888
1877 Pecten trifidus Dunker, 1877
1882 Pecten jickelii Dunker, 1882 [nomen novum pro Pecten trifidus Dunker, 1877]
1888 Pecten hysginoides Melvill, 1888

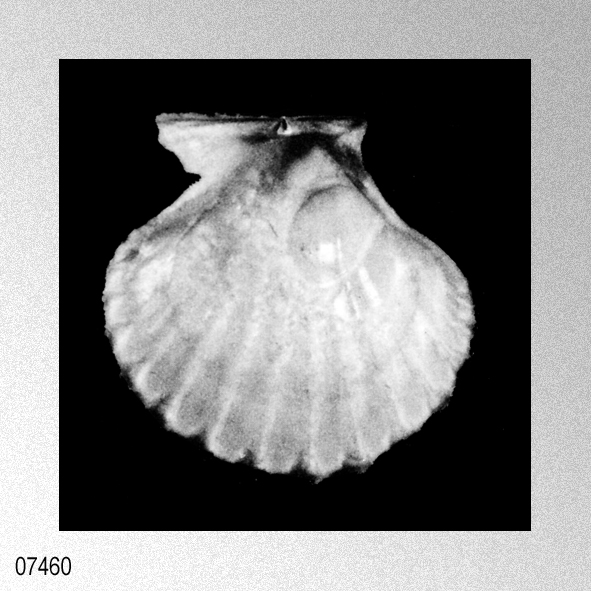
Pecten vesiculosus Dkr.; W. Dunker, 1882, Index Molluscorum maris Japonici, plate 11, figure 1.
|
«Testa solidula paullum obliqua inaequivalvis costis radiantibus 15 aequalibus rotundatis obsolete striatis paene laevibus, utrinque lamellatis, interstitiis tenerrime striatis transversimque cancellatis instructa; auriculae impares acute costatae, ad marginem cardinis utrinque plicati subserratae; color externus rubescens vel albus rubroque variegatus, internus albus et roseus. — Alt. spec. maximi 27 mill.»
WILHELM DUNKER, 1877
|


















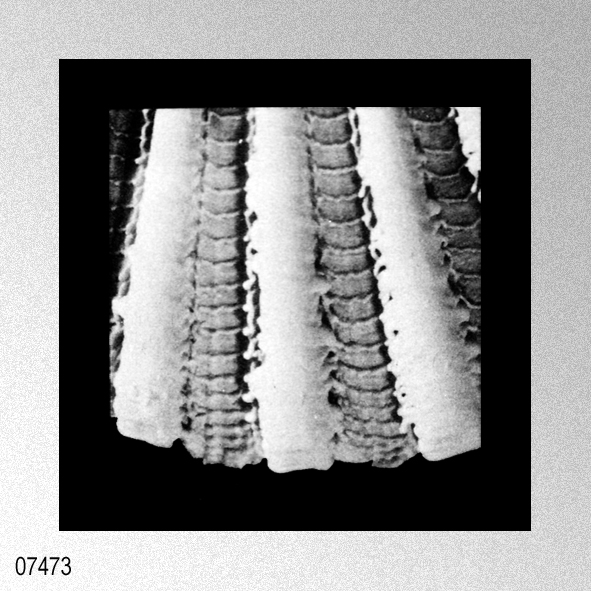



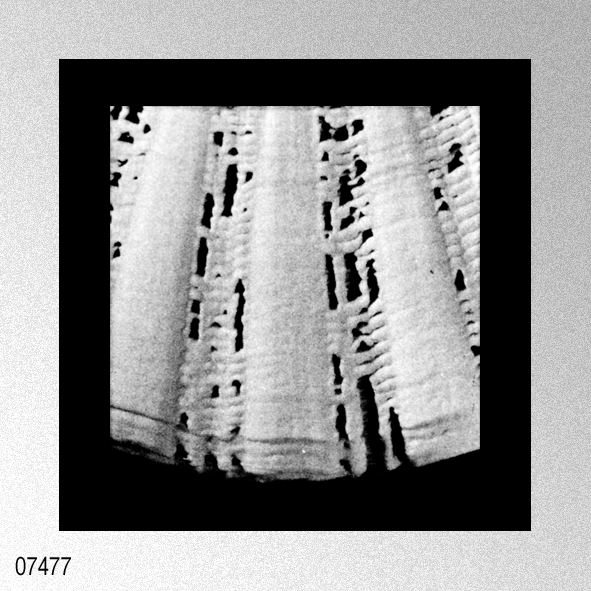


























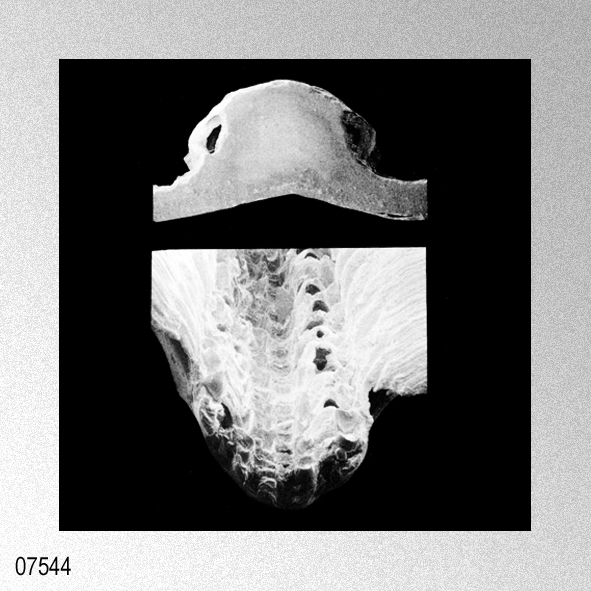
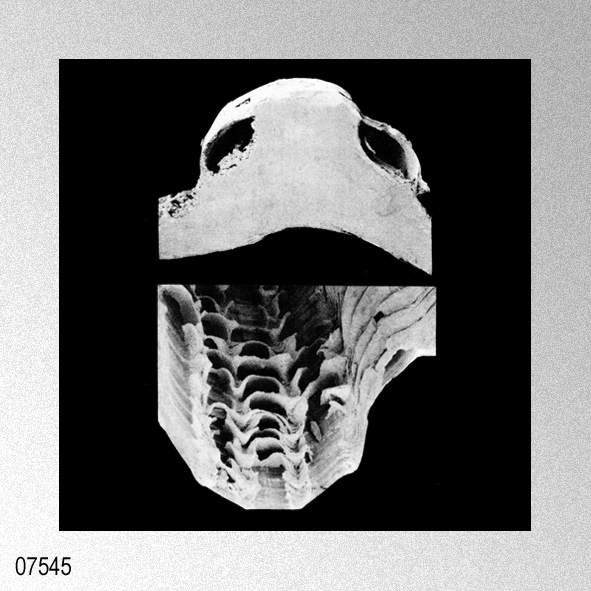

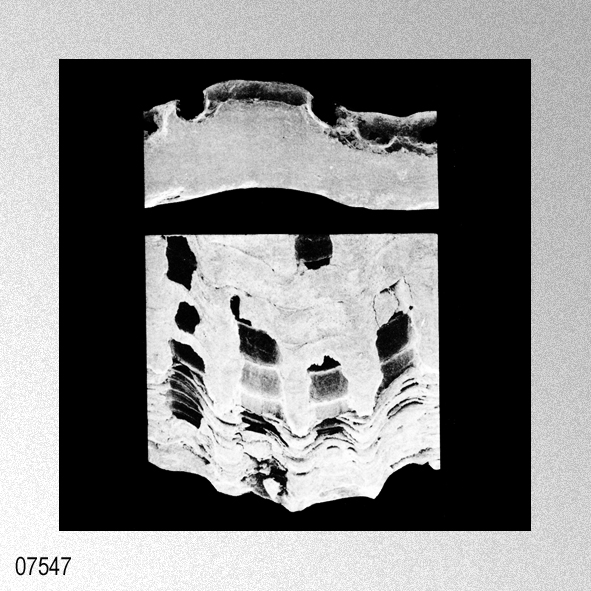



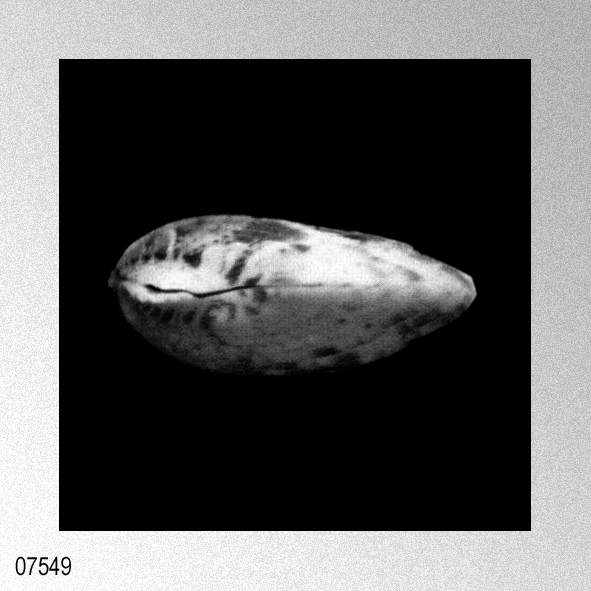
|
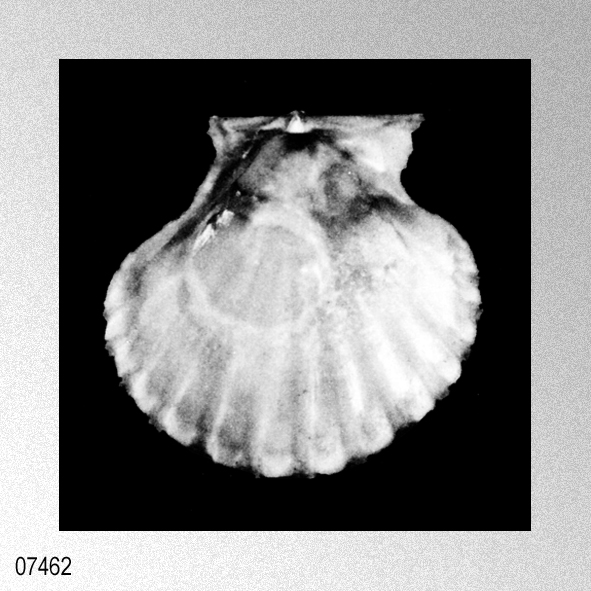
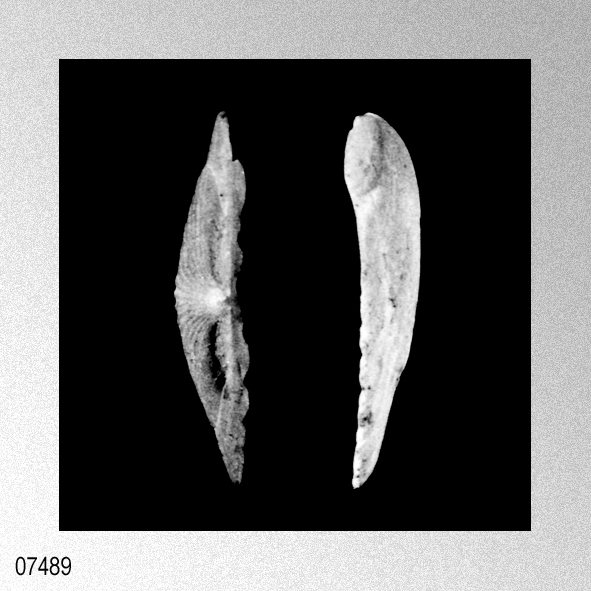
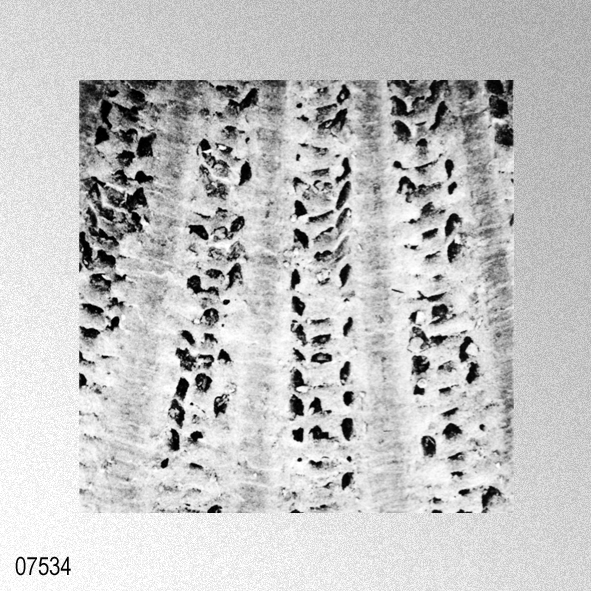
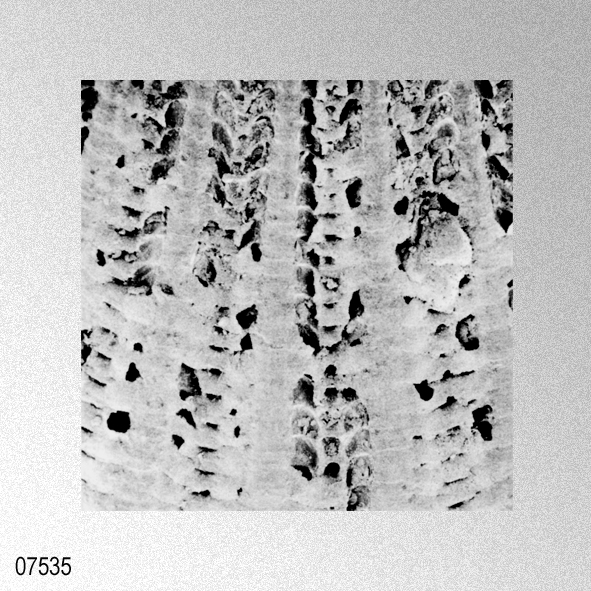
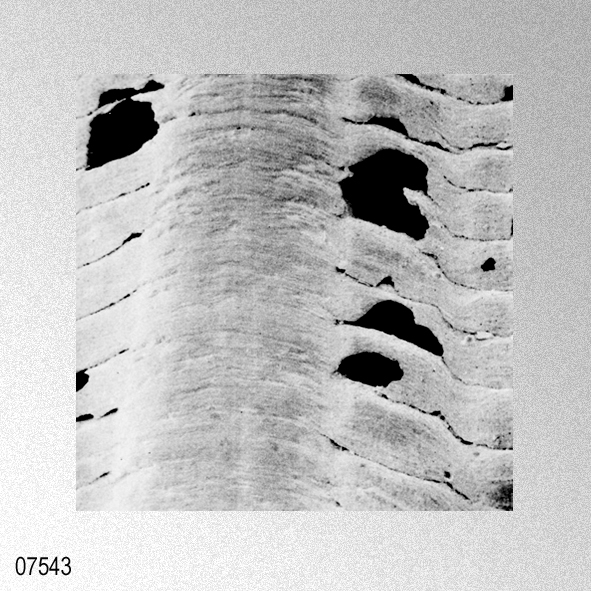
«Description.— Shell solid, oblong, somewhat wider than long, max. h. 29.9 mm, w. 30.6 mm, d. 15.5 mm. Valves inaequilateral, right valve up to 1.6 times more convex than left one; lunulae smooth; left valve with 16-17 equally pronounced ribs; right valve with 16-17 ribs, also equally pronounced. Microsculpture as described for the genus, with the sole exception in this species that the central wall connecting the two alternating rows of vesicles is rather thick, this in contrast to the other species discussed. Anterior auricles 1.5-1.6 times as large as the posterior ones; anterior auricle of left valve with 5-6 ribs, of right valve with 6; posterior auricle of left valve with 5-6 ribs, of right valve with 6-7. Byssal notch broad and short, 3-5 teeth observed. Resilium small and triangular. Top angle 95°-99°. Colour pattern of the shell generally consisting of brown, reddish or violet with white or cream V-shaped markings.
Remarks. — This species is much more solid than any of its congeners, and often has between 16 and 22 mm from the apex a strong growth line, which results in larger specimens in so-called box-shaped forms. This phenomenon was not encountered in the other species discussed. Distribution.— ln contrast to the other taxa this species seems to be restricted to central and southwest Japan and adjacent seas.» WAGNER, H. P. 1989. The genus Cryptopecten Dall, Bartsch & Rehder, 1938, in the Indo-Pacific (Mollusca; Bivalvia; Pectinidae). Basteria, 53: 53-62, figs. 1-18. [p. 55]
|

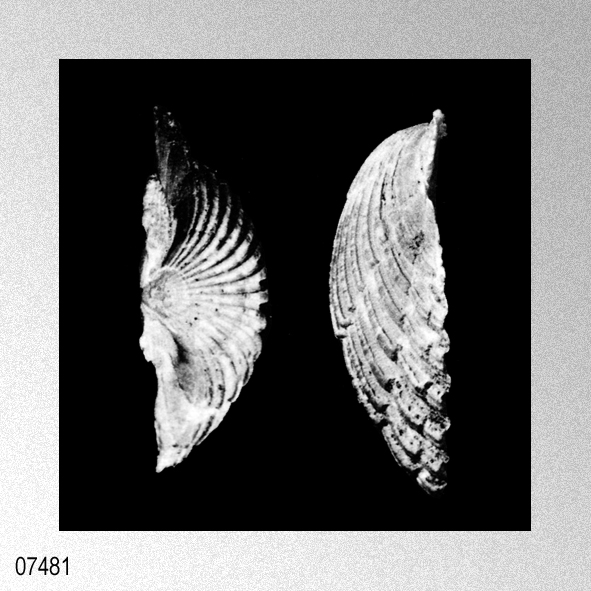
Cryptopecten vesiculosus (Dunker, 1877); H. P. Wagner, 1989, The genus Cryptopecten Dall, Bartsch & Rehder, 1938, in the Indo-Pacific, figures 1 (holotype Pecten hysginoides Melvill), 2.
|
|
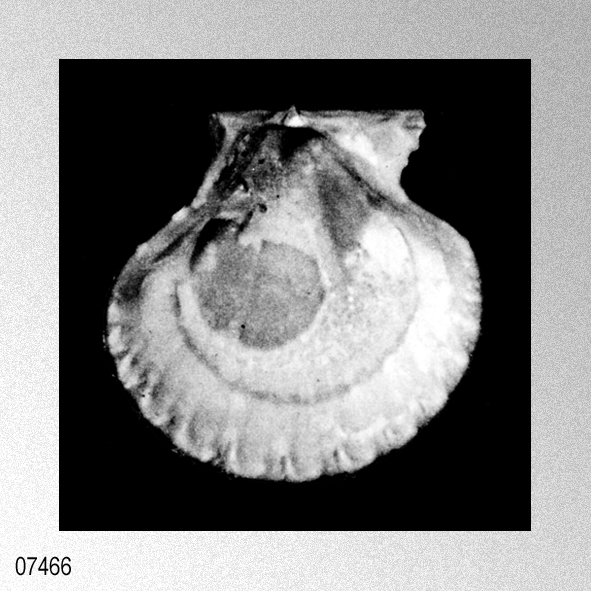
«Seven specimens are assigned to this species on account of their moderately thick, suborbicular, inequilateral shell which has 15 to 17 radial ribs. Each radial rib consists of a central solid ridge and hollow lateral parts which are ornamented with oppositely disposed imbricated scales. The morphological description and intrapopulational variation of this species were fully treated by Hayami ( 1973, 1982, 1984 ). The species is known to live in the sublittoral zone in warm waters under the influence of the Kuroshio current and its branch around the Japanese Islands and the South China Sea, and has also been reported in shallow water assemblages of warm and temperate conditions of Pliocene and Pleistocene time. This record is one of the northernmost occurrences of the present species in the Late Pliocene.»
O'HARA, S. & N. NEMOTO. 1988. Pectinids from the Taga Group of the Joban Coalfield. Saito Ho-on Kai Special Publication, Professor Tamio Kotaka Commemorative Volume: 481-496, pls. 1-4. [p. 489, 490]
|

Cryptopecten vesiculosus (Dunker, 1877); S. O'Hara & N. Nemoto, 1988, Pectinids from the Taga Group of the Joban Coalfield, plate 2, figures 1, 3.
|
|
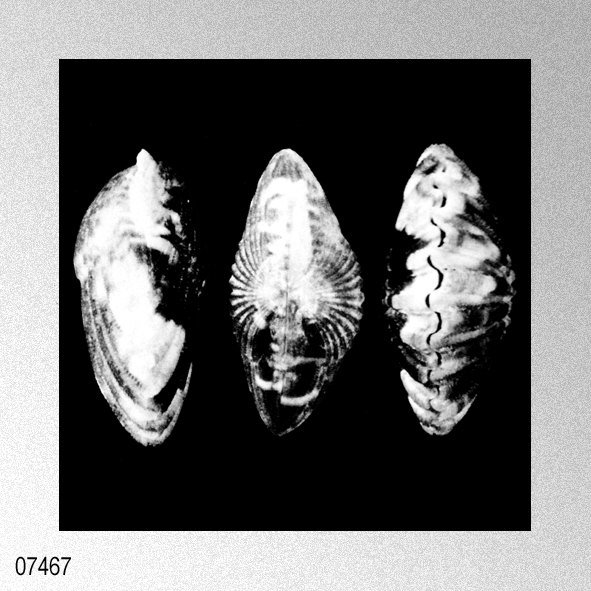
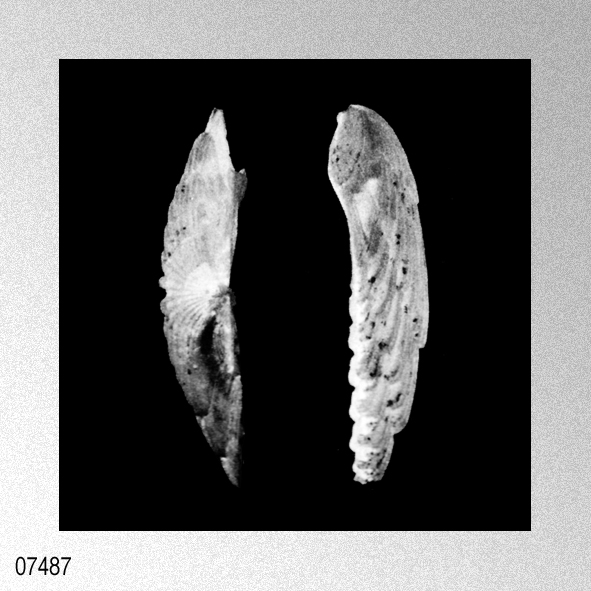
«Remarks.— Cryptopecten vesiculosus is a common pectinid in Japan, but its morphology and variation have not been studied in detail. The remarkable discontinuous variation of disk sculpture, however, seems to have been noticed by some previous authors. For example, Kuroda (1932) illustrated two left valves belonging to different phenotypes in juxtaposition. Furthermore, Habe and Kosuge (1967) and Kuroda, Habe and Oyama in Biological Laboratory, Imperial Household (1971) mentioned, though briefly, the presence of dimorphic phenomenon in their materials. Fortunately, however, except for Kira (1950) and Mori and Osada (1979), no one seems to have regarded the two phenotypes as separable into different taxa. As already documented in this paper, the two phenotypes coexist in every large sample after the Middle Pleistocene and are surely attributable to intrapopulational discontinuous variation.
Some problems about the synonymy of early proposed specific names remain unsolved. Pecten trifidus Dunker, 1877, from Japan, which was renamed Pecten jickelii by Dunker (1882) without any adequate reason, may be conspecific with P. vesiculosus, as was suggested by Habe (1977). The original descriptions of P. trifidus and P. jickelii were not accompanied by any illustrations, but the number of radial ribs, which was said to be 18 in P. trifidus, may be somewhat larger than the averages in the present samples of C. vesiculosus. Their taxonomic position, therefore, is difficult to determine without examining the type material. On the basis of my observation of Dr. T. R. Waller's unpublished photographs, I conclude that the type conjoined specimen of Pecten hysginodes Melvill, 1888, from unknown locality, which is preserved in the Cardiff Museum, may belong to the Phenotype R of C. vesiculosus. The detailed morphology and phenotypic frequency are, as described in the preceding chapters, considerably variable in time and space, but all the samples here treated certainly belong to a single evolutionary species. Masuda (1962) mentioned the existence of some morphological difference between the Pliocene and Pleistocene-Recent specimens of this species. He pointed out that some intercostal threads are developed in the Recent and Pleistocene specimens near the ventral margin, but that they are usually absent in Tertiary ones. Furthermore, the Pliocene specimens were said to have broader ribs and narrower interspaces than Pleistocene-Recent ones. My observation of the present samples of various ages revealed these characters to be more closely related to phenotypic and subphenotypic differences, ontogenetical change and state of preservation than to chronological change. The intercostal sculpture actually differs not only between the two phenotypes but also between the two subphenotypes (p. 58) within Phenotype Q; intercostal threads commonly appear in adult individuals of rough subphenotype but are rarely met with in those of smooth subphenotype almost regardless of geological horizons. Many well-preserved large specimens from the Pliocene, for example, those in samples Sh 1 and Ik, show as clear intercostal threads as those of Pleistocene-Recent samples, while smaller specimens always lack such threads. The relative breadth of radial ribs varies considerably among individuals within one sample and increases remarkably with growth. A striking feature is that radial ribs appear very narrow if imbricated scales on their sides are exfoliated. Though biometrical comparison of this character among samples has not been carried out orving to the different growth stages and difficulty of subphenotypic discrimination in many unfavorably preserved samples, chronological change could not be detected with respect to the relative breadth of ribs and interspaces. Distribution.— This subspecies lives on the outer sublittoral sandy substrates around the central and southwestern parts of the Japanese Islands under the influence of Kuroshio warm current (35°N and south) and its branch, Tsushima Current (41°N and south) (see Figs. 1 and 3). Fossils of this subspecies are also almost entirely restricted to the same area as the distribution in the present time, though the northern limit seems to have fluctuated with time (see Fig. 4). Middle Pliocene (ca. 3.5 Ma) to Recent.» HAYAMI, I. 1984. Natural history and evolution of Cryptopecten (A Cenozoic-Recent Pectinid Genus). The University Museum, The University of Tokyo, Bulletin, 24: 1-149, pls. 1-13. [p. 108]
|

Cryptopecten vesiculosus vesiculosus (Dunker); I. Hayami, 1984, Natural history and evolution of Cryptopecten, plate 6, figures 1-6.
|
|
|


    |