Spondylopecten Roeder 1882
ROEDER, H. A. 1882. Beitrag zur Kenntnis des Terrain à Chailles und seiner Zweischaler in der Umgegend von Pfirt im Ober-Elsass. Schulz & Co., Strasburg
|
«Das interessanteste an diesen Formen ist jedenfalls das Schloß, und darüber geben meine verkieselten Exemplare guten Aufschluß. Auf der rechten Klappe sieht man unter dem wenig gewölbten Wirbel eine verhältnismäßig breite Area, die durch die Bandgrube wie bei Spondylus gespalten ist.
Neben der Bandgrube liegen unter der Area jederseits Zähne und zwar ist der vordere bedeutend größer, vorausgesetzt, daß der hintere nicht teilweise abgebrochen ist, was bei meinen Exemplaren immerhin möglich wäre. QUENSTEDT hebt diese Ungleichheit der Zähne jedoch auch für seinen Pecten globosus .... auf den ich gleich noch zu sprechen komme, hervor, deshalb scheint sie also normal zu sein. Der große Vorderzahn ist löffeiförmig in die Höhe gekrümmt und zeigt auf der Area zugekehrten Seite senkrechte, parallel Streifen; der Hinterzahn ist klein und undeutlich, er erhebt sich kaum über die Area. Außerdem ist der gerade Schloßrand, vorn der Oberrand des Ohres, mit feinen senkrechten Kerben versehen. Von der linken Klappe besitze ich leider kein ganz erhaltenes Schloß; ich habe nur die senkrechten Kerben auf dem Schloßrande constatiren können.' (relevant extract from description of Pecten (Spondylopecten) cf. erinaceus BUVIGNIER; ROEDER, 1882).» H. A. ROEDER, 1882
[from A. L. A. Johnson, 1984, The palaeobiology of the bivalve families Pectinidae and Propeamussiidae in the Jurassic of Europe, p. 90, 91] |

Pecten erinaceus, Buv.; A. Buvignier, 1852, Statistique géologique, minéralogique, minérallurgique et paléontologique du Département de la Meuse, plate 19, figures 7-12.
|
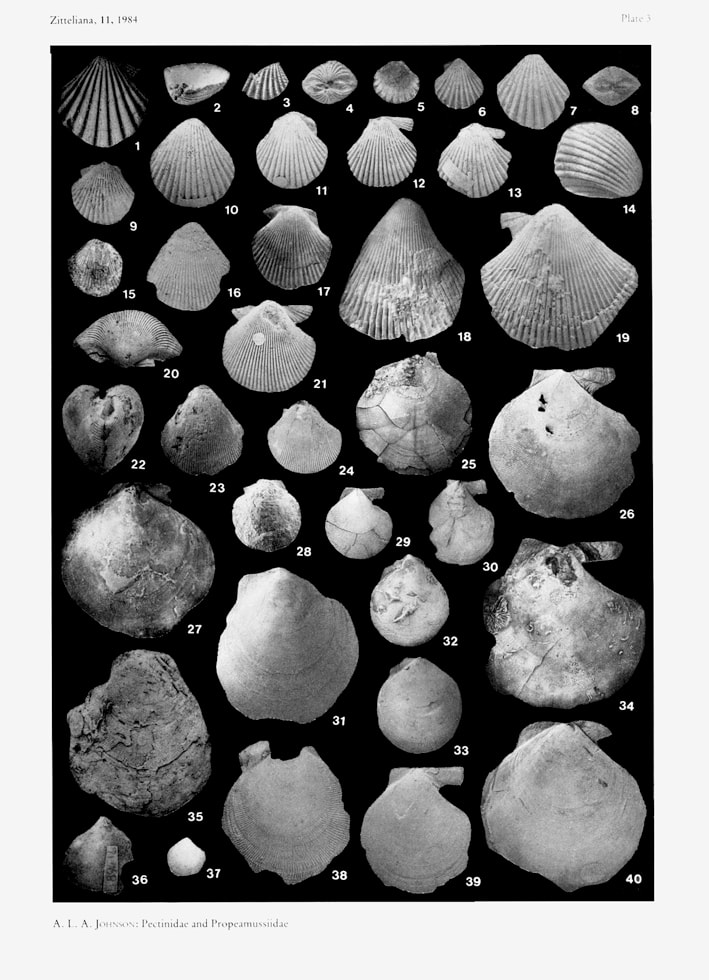
Spondylopecten (Spondylopecten) palinurus (d'Orbigny 1850); A. L. A. Johnson, 1984, The palaeobiology of the bivalve families Pectinidae and Propeamussiidae in the Jurassic of Europe, plate 3, figures 8-14.
|
«AMENDED DIAGNOSIS
Spondylopecten with rounded plicae bearing 2-4 rows of spines. Jur. (Aalen. - U. Tithon.), Eu., Afr., Asia.
DISCUSSION
In general form all members of S. (Spondylopecten) are very similar. It is possible therefore that the sub-groups identified below could be polymorphs of the same species. However, with the apparent impossibility of detecting polymorphism in the fossil record it seems preferable to treat them as separate species.
In measured museum specimens the ränge of plical variation in S. (Spondylopecten) is from 18-71. Within the ränge 18-36 plicae there is an essentially bimodal distribution grouped around modes at 21 and 30/32 plicae with an intervening trough at 26 plicae (text fig. 75). Individuais in the former group (herein referred to S. (S.) palinurus) also seem to differ from those in the latter (herein referred to S. (S.) subpunctatus) by their lower convexity (text figs. 77, 84) and in the possession of four rather than two rows of plical spines. The single known specimen with 26 plicae (MNP) is abraded but apparently originally bore four rows of spines. It is therefore considered to indicate the upper limit of plical Variation in the former group. Within the first group later populations differ in the mode and ränge of plical variation (text fig. 79) and in convexity (text fig. 77). While there can be little doubt of an ancestor descendant relationship some authors would consider such differences worthy of a specific Separation. However, apart from the difficulties of obiectively defining the species there is no evidence that separate lineages existed at any one time. In fact there is some evidence (see p. 97) that earlier and later populations were linked by gradual phyletic evolution. Thus following the rationale adopted in this work, the earlier and later populations are herein considered to belong to the same species. Similar reasoning can be applied to the group of forms referred to S. (S.) subpunctatus in which later samples differ in the mode and ränge of plical Variation (text fig. 86) and in convexity (text fig. 84) but in which there is no evidence for coexisting lineages, yet a certain amount for phyletic gradualism (see p. 102). The paucity of available museum specimens renders it difficult to make any objective division in forms with more plicae than S. (S.) subpunctatus. Except where indicated, museum specimens with the following plical counts are only known singly: 42, 43 (3), 44 (3), 45, 46, 48, 51, 52, 53 (2), 54, 56, 60, 63, 64, 71. However, foreign authors who have undoubtedly had access to a greater volume of material provide good evidence for a bimodal distribution in the frequency of plical counts. STAESCHE (1926) refers to specimens with about 45 plicae while YIN (1931) refers to a number of specimens with bet\wen 42 and 48 plicae, thus forms within the latter range are considered to belong to a third S. (Spondylopecten) species, (S. (S.) cardinatus). BOEHM (1883) records 55 specimens wilh over 60 plicae while STAESCHE (1926) records numerous specimens with between 55 and 65 plicae, thus forms within lhe latter range, together with rare museum specimens with plical counts just outside this range (51, 52, 53, 54, 71) are considered to belong to a fourth S. (Spondylopecten) specIes (S. (S.) globosus).» JOHNSON, A. L. A. 1984. The palaeobiology of the bivalve families Pectinidae and Propeamussiidae in the Jurassic of Europe. Zitteliana, 11: 1-235, pls. 1-11. [p. 91, 92]
|