Chlamys marshallensis Waller, 1972
WALLER, T. R. 1972 The Pectinidae (Mollusca: Bivalvia) of Eniwetock Atoll, Marshall Islands. The Veliger, 14 (3): 221-264, pls. 1-8, text figs. 1-22. [p. 236, pl. 2, figs. 26-33, 36-37; text figs. 3, 16]
|
Chlamys marshallensis Waller, spec. nov.
(Figures 26 - 33, 36, 37; Figures 3, 16; Table 4) Types
«The holotype of Chlamys marshallensis is a complete shell with soft parts separately preserved in alcohol and bearing the catalogue number USNM 701200. The type locality is the lagoon of Eniwetok Atoll at 71°24'12'' N; 162°22' a coral knoll at a depth of 65 feet. At this locality (on Sepcoral knoll at a depth of 65 feet. At this locality (on September 27, 1969) the author, using SCUBA equipment, found the specimen living deep within a crevice in a head of Porites. The height (AM) and length (AG) of the holotype are 13.5 and 11.9 mm, respectively. Paratypes are those specimens listed below under "Specimens examined."
Description
Shell outline: Shell small in size, not known to exceed 30 mm in height, commonly about 20 mm; somewhat higher than long and approximately equiconvex. Valves somewhat flattened near ventral margin, generally with commissure somewhat distorted. Left valve very slightly shorter in height than and fitting within ventral margin of right valve. Outline of disk of mature individuals oblique and somewhat prosocline.
Auricles and outer ligament: Right anterior auricle with dorsal margin on crest of fold that increases in amplitude distally; byssal notch deep, with pointed or curved apex floored by ctenolium with 6 to 8 teeth; byssal fasciole broad and slightly concave. Left anterior auricle with ventral portion curved inward through plane of commissure, extending slightly into byssal notch of right valve. Posterior auricles much smaller than anterior, with dorsal margins slopinq ventrally in distal direction and free margins straight or somewhat concave, forming obtuse angles with dorsal margins. Length of anterior outer ligament about 1½ times that of posterior outer ligament; entire outer ligament migrating ventrally (extent of migration up to 0.8 mm from origin of growth in individuals 16 to 22 mm in height), leaving band of ligament material that tapers to distal extremities.
Exterior shell surface: Mature valves with fine rounded, flattened, or broadly angular radial costae appearing at heights from 4.5 to 8.5 mm on right valves and from 8.5 to 14 mm on left valves (measured in central area of disk); primary costae closely spaced on lateral areas of disk and becoming scabrous early; in central area more wideiy spaced, becoming scabrous near ventral margin, and on some individuals becoming separated by from 3 to 5 very low, flattened, non-scabrous secondary costae that may also become scabrous at distal edge of valve. Height of exterior prismatic layer on right valve 0.84 to 0.92 mm; Camptonectes microsculpture dominant feature on surface of both valves before start of primary radial costae; Camptonectes striae in central area of disk aligning with and merging into early secondary costae. Concentric lamellae extremely faint and not projecting, most readily discernible on distal half of right valve. Anterior auricle of mature individuals with 4 to 6 costae distally on right valve, 6 to 11 on left; 2 to 6 costae on each posterior auricle. Disk flanks smooth or with incipient costae distally. Color of shell highly variable, commonly white with faint maroon rays or radially elongate spots; less commonly uniform lemon yellow, pale orange, or coral red, or with colors arranged in polygonal spots formed by a pattern of intersecting diverging lines curuing ventrally and laterally from the beak. Pattern of intersecting opaque white lines common on first few millimeters of growth of disk of right valve.
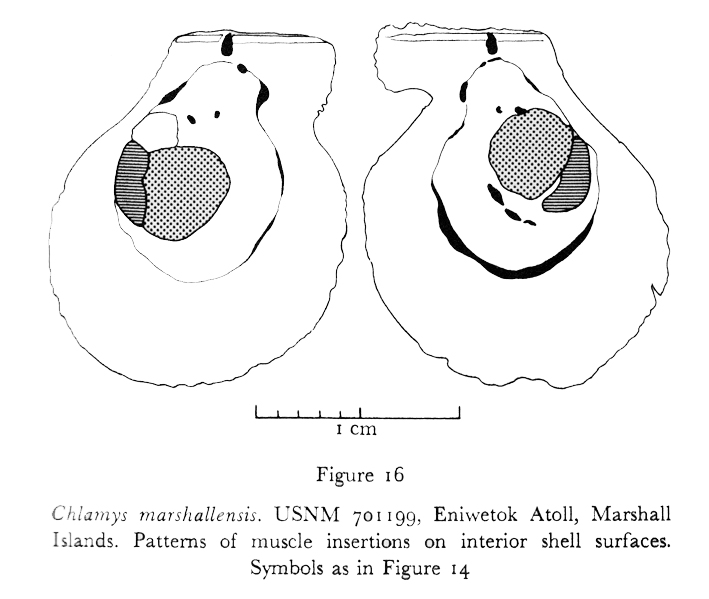
Internal features: Resilial insertions very high and narrow due to ventral migration of active ligament area; acline to rather strongly prosocline. Pattems of muscle insertions on valve interiors as in Figure 16. Vela translucent, either uniformly dark in color (commonly maroon) or more commonly with broad, vertical bands of altemating light and dark color, generally shades of brown. Fringing tentacles commonly longitudinally striped, not clearly distinct from exploratory tentacles; exploratory tentacles tending to be in groups, either colorless or with narrow pigmented rings, separated by groups of fringing tentacles that are darker in color; guard tentacles either uniformly translucent or with small rings of granular pigment. Eye color orange-red.
Comparison
Chlamys marshallensis most closely resembles Chlamys squamosa (Gmelin, 1791), an apparently allopatric species that is widely distributed in the western pacific (Figures 3, 38 - 41; Table 5). The new species differs from C. squamosa in having primary costae that begin later. The first clearly continuous primary costae appear at heights 4.5 to 8.5 mm on right valves and 8.5 to 14 mm on left valves of C. marshallensis. Chlamys squamosa generally has right-valve primary costae beginning at the edge of the prismatic layer, about 1 mm from the origin of growth, and left-valve primary costae beginning well up on the beak. In addition, C. marshallensis differs in having much more prominent Camptonectes microsculpture on both valves and secondary costae in the center of the disk that begin earlier through the radial alignment of Camptonectes striae. Both the primary and secondary costae of mature C. marshallensis become scabrous, whereas at the distal edge of C. squamosa broad scabrous primaries are separated by from 1 to 5 smooth secondary costae in the central portion of the ventral margin. The maximum size reached by C. squamosa, 50 to 60 mm, exceeds the known maximum of about 30 mm for C. marshallensis.
The new species also resembles Chlamys sp. cf C. irregularis (G. B. Sowerby 2nd, 1847), a species of about the same size and coloration that has thus far been found in the Cocos-Keeling, Caroline, Marshall, Gilbert, and Phoenix Islands and possibly also in Samoa (Figures 3, 34, 35). If conspecific with true C. irregularis, this species also occurs in Japan (Hirado; Kii Peninsula). Chlamys sp. cf. C. irregularis differs from C. marshallensis in having primary costae that extend to the prismatic border on its right valve and well onto the beak of its left valve; secondary costae that are higher and also begin earlier, interrupting the Camptonectes microsculpture; and right-valve costae that all becorne scabrous before a height of about 13 mm is reached. Chlamys madreporarum (G. B. Sowerby 2nd, 1847), a species of Japanese and Malaysian waters that apparently lives deep within corals in a manner somewhat like Pedum (see ecology of Pedum below), differs from C. marshallensis in having a more oblique form with greatly elongated posterior slope and more closely spaced and fasciculatecl radial costae that are closely scabrous and extend well up into the umbonal areas (Figure 42). The Hawaiian species Chlamys cookei Dall, Bartsch, & Rehder, 1938, shown in Figures 43 and 44, differs from C. marshallensis in having more numerous primary costae. Chlamys midwayensis Habe & Okutani, 1968, is a possible synonym of C. cookei. Functional Morphology
Chlamys marshallensis, like other Chlamys that live byssally attached deep in coral crevices (e. g., C. madreporarum and C. irregularis), develops a thin, fragile shell commonly elongated in a postero-ventral direction and having a disproportionately large pair of anterior auricles. Postero-ventral elongation appears to result from a physical restriction of growth in other distal directions by the nearly enclosing coral.
It has been observed that C. marshallensis, when forcibly detached in its natural habitat, can move laterally in search of a new attachment site. Movement appears to be distinctly toward dark areas. Using its thin shell of low convexity unencumbered by strong plication or surface rugosity, the scallop can slide into a narrow crevice. Then through propulsion by clapping its valves followed by repeated sequences of foot extension, byssal secretion, pedal contraction, and byssal release, the animal wedges itself tightly deep within the crevice. The ventralty migrating outer ligament and hinge line (Figures 36, 37) allow the animal to continue to open its valve even though its size is increasing within a confined space, in the same manner as that described for Hinnites and Pedum by Yonge (1951, p. 416; 1967, p. 321). The long right anterior auricle of byssally attached Chlamys appears to serve as a lever arm to allow the scallop to pull down tightly over its byssal attachment against a flat surface without rotating the shell into a position that is nearly vertical to the surface of attachment (STANLEY, 1970, p. 31). This functional interpretation is supported by the orientation of the hinge line relative to the plane of commissure (Figures 27, 31). In most byssally attached true Chlamys, it can be seen in dorsal view with the right valve downward and the plane of commissure horizontal that the hinge line slopes downward from posterior to anterior. This slope, together with the strong dorsal fold of the right antenior auricle, allow the valves to be maintained in a position more nearly parallel to the attachment surface. Ecology
Living habits: Chlamys marshallensis appears to favor lagoons rather than outer reefs. In the Eniwetok lagoon it lives out of sight pulled tightly by byssal atachment against branches of live Acropora, the lower surfaces of
dead palmate Acropora, or the concave under surfaces of massive heads of Porites. It was also found byssally attached deep within crevices in heads of Porites. When forcibly detached the species swims rapidly with very rapid adductor contractions, rising from the bottom at rather steep angle and moving laterally up to about a foot at an altitude from several inches to a loot from the bottom. In spite of the oblique, asymmetrical shape of the shell, direction of movement seems to be fairly straight, without spiraling. Depth records of live specimens: Eniwetok Lagoon, 15 to 65 feet; Bikini Lagoon, 90 to 180 feet; Rongelap Lagoon, 120 feet; Arno Lagoon, "on lagoon reef and slope."
Geographic distribution: Known only from the Marshall Islands (Eniwetok, Bikini, Rongelap, Kwajalein, and Arno Atolls; see Figure 3).
Fossil Records
No fossil representatives of Chlamys marshallensis have been found in the collections of the U. S. National Museum or in the illustrated literature.
Specimens Examined
Chlamys marshallensis
Marshall Islands: ENIWETOK ATOLL: lagoon off S end of Chimini Island, 45 feet, byssally attached among branching coral (USNM, live); reef in lagoon between Chinieero Island and Van Island, 15 - 20 feet, byssally attached below ledges of dead Acropora (USNM, live); Rigili Island, 25 feet, reef flat in lagoon (live), specimen lost in transit; off Parry Island, 65 feet, in lagoon on coral pinnacle, deep within crevice on head of Porites lutea and clinging to concave undersurface of head (USNM 701200, holotype, live and dead ); Eniwetok Lagoon, 50 feet (ANSP, live) ; BIKINI ATOLL: ¼ mile S of W end of Bikini Island, 15 fathoms in lagoon (USNM, live); 4 miles S of W end of Bikini Island, 28 fathoms (USNM, live); 4 miles S of W end of Bikini Island, 25 fathoms, in lagoon (USNM, live); Bikini Lagoon, 25- 30 fathoms (USNM, live); RONGELAP ATOLL: 20 fathoms, in lagoon (USNM, live); KAWJALEIN ATOLL: beach drift (USNM, dead) ; Loi and South Loi Islands, beach drift (USNM, dead ); ARNO ATOLL: Ine Island, near edge of lagoon reef on coral, Ceriatopora hystrix (USNM, probably live); Ine Island, outer lagoon slope and reef flat, in porous rock (USNM, probably live); Ine Island, outer slope of lagoon reef flat, in Acropora heads (USNM, live and dead); Ine Island, on lagoon reef (USNM, dead).
Chlamys squamosa
Malaysia, Thailand: (USNM, live). Japan: Osumi, Okinawa, Ryukyus (USNM, live and dead). Philippines: (USNM, ANSP, live and dead). Australia: (ANSP, live). Dutch New Guinea: (USNM, ANSP, live and dead). New Caledonia: (ANSP, live and dead). Fiji Islands: (USNM, live and dead). Mariana Islands: (USNM, ANSP, live). Caroline Islands: (USNM, ANSP, Iive and dead). Samoa: (UNSM, ANSP, live). Tonga Islands: (USNM, live).
Chlamys sp. cf. C. irregularis
Japan: Okinawa (ANSP, live). Caroline Islands: IFALUK ATOLL: Rauau village, Rolong, Falarik Island, from Porites reef in lagoon over muddy turtle grass flats (USNM, live); Falarik Island, pass in coral (USNM, live); Katelu benju, Rauau, Falarik Island, lagoon, in Pocillopora (USNM, live). Marshall Islands: KAWAJALEIN ATOLL: beach drift (USNM, dead). Gilbert Islands: ONOTOA ATOLL; about 3¼ miles N 30 W, from Tabuarorae Maneaba, near the center of Te Rawa ni Bao, a pass in the S part of the leeward reef from thickly set coral masses (USNM, live). Phoenix Islands: near Flemings, inside coral in lagoon (USNM, live ). Samoa: Assau Harbor, N coast of Savaii, along causeway (USNM, live).»
THOMAS RICHARD WALLER, 1972
|

T. R. Waller, 1972, plate 2.
|